Download
1 / 63
630 likes | 924 Views
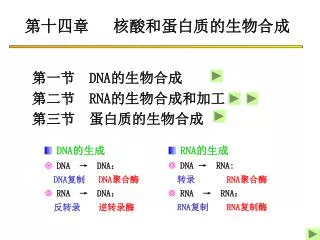
第十四章 核酸和蛋白质的生物合成. 第一节 DNA 的生物合成 第二节 RNA 的生物合成和加工 第三节 蛋白质的生物合成. RNA 的生成 DNA → RNA: 转录 RNA 聚合酶 RNA → RNA : RNA 复制 RNA 复制酶. DNA 的生成 DNA → DNA : DNA 复制 DNA 聚合酶 RNA → DNA : 反转录 逆转录酶. 第一节 DNA 的生物合成. 一、 DNA 的复制 二、 RNA 的逆转录. 一、 DNA 的复制. 半保留复制 复制子 半不连续复制

E N D
第十四章 核酸和蛋白质的生物合成 第一节 DNA的生物合成 第二节 RNA的生物合成和加工 第三节 蛋白质的生物合成 • RNA的生成 • DNA → RNA: 转录RNA聚合酶 • RNA → RNA: RNA复制 RNA复制酶 • DNA的生成 • DNA → DNA: DNA复制 DNA聚合酶 • RNA → DNA: 反转录 逆转录酶
第一节 DNA的生物合成 一、DNA的复制 二、RNA的逆转录
一、DNA的复制 • 半保留复制 • 复制子 • 半不连续复制 • DNA复制体系 • DNA复制过程
(二) 复制子 P408 • 复制子:基因组中能单独进行复制的单位。每个起始点到终止点的区域为一个复制子。 • 单复制子: 一般原核生物DNA • 多复制子:真核生物染色体DNA,具有多个复制起点。 • 起始点(origin of replication, ori, O)(复制原点): 复制子中控制复制起始的位点,富含AT。 • 终止点:终止复制的位点。 • 复制的方向:单向或双向 (5 ´→ 3´)
单向复制 双向复制 复制叉(生长点)
(三) DNA的半不连续复制 • 冈崎片段 前导链 滞后链 引物RNA
(四)DNA复制体系 413页 • 底物: dNTP (dATP, dGTP, dCTP ,dTTP) • 模板: 单链的DNA • 引物: 寡核苷酸引物(RNA) • 酶和蛋白质因子
DNA复制中出现的酶 • DNA聚合酶 • 以4种dNTP为底物 • 反应需要接受DNA模板的指导,产物DNA的性质与模板相同 • 反应需有引物 3 ’-OH的核酸链,不能催化游离的dNTP的聚合。 • 链生长方向 5’→ 3 ’ • DNA连接酶(ligase) • 解螺旋酶(helicase) • DNA旋转酶(拓扑异构酶) • 单链结合蛋白 • 引物合成酶(引发酶,RNA聚合酶,primase)
DNA聚合酶 P414 大肠杆菌中含有5种不同的DNA聚合酶 • DNA聚合酶Ⅰ • DNA聚合酶Ⅱ • DNA聚合酶Ⅲ • DNA聚合酶Ⅳ • DNA聚合酶Ⅴ 涉及DNA的错误倾向修复。当DNA受到严重损伤的时候,就可以诱导产生这两个酶,使修复缺乏准确性。出现高突变率。
5 3 ´核酸外切酶 3 5 ´核酸外切酶 聚合酶 作用于双链DNA的碱基配对部分,从5 ´端来水解寡核苷酸 DNA聚合酶Ⅰ 不是复制酶,而是修复酶 • 具有DNA聚合酶活性,可以使DNA链沿5´→3´方向延长, • 它还具有3´→ 5´核酸外切酶的活性,由3´端开始水解DNA链; • 它具有它还具有5´→3´核酸外切酶的活性,由5´端开始水解DNA链; 切除单链DNA的3 ´末端核苷酸,对双链DNA不起作用 双重校对
DNA聚合酶Ⅱ • DNA聚合酶Ⅱ是多亚基酶 • 以dNTP为引物,从5´→ 3´方向合成DNA • 带有3 ´→ 5 ´外切酶的活力,但是没有5´→ 3´核酸外切酶的活力 • 也是一种修复酶,而不是一种复制酶 DNA聚合酶Ⅲ • 是一个多亚基的酶(核心酶为αεθ) • 是DNA的复制酶 • 在模板的指导下,以dNTP为底物,按照5 ´→ 3´方向聚合DNA ,具有3´→5´外切核酸酶的活性,但是没有5´→3´核酸外切酶的活性。
(五) DNA复制过程 P421 1.大肠杆菌 起始阶段 复制的延伸 复制的终止
2 真核生物 • 与原核生物复制的区别: • 真核生物染色体有多个复制起点 • 真核生物的复制速度要慢于原核生物 • 真核生物染色体在全部复制完成之前不再从新开始复制。 • DNA聚合酶α、β、γ、δ、ε
二、RNA的逆转录(RNADNA) P493 • 以RNA为模板合成DNA的过程叫做逆转录。 • 逆转录酶(reverse transcriptase, RT)是个多功能酶: • 逆转录活性: cDNA:合成出的相应于一定mRNA的互补DNA。 • RNase H 活性: • DNA聚合酶活性: • 常用的逆转录酶有两种: • 来自鸟类成髓细胞瘤逆转录病毒称为AMV-RT; • 来自鼠白血病毒称为MMLV-RT。 • RT的发现对于遗传工程技术起了很大的推动作用,它已成为一种重要的工具酶。用组织细胞提取mRNA,在反转录酶的作用下,合成出cDNA,由此可构建出cDNA文库,从中筛选特异的目的基因,这是在基因工程技术中最常用的获得目的基因的方法。
第二节 RNA的生物合成和加工 • RNA复制 • 转录
病毒RNA 组装成新病毒 复制 RNA复制酶 互补RNA链 复制 新病毒RNA 翻译 病毒蛋白 一、RNA的复制 • RNA复制酶 • 以病毒RNA做模板,再由4种核苷三磷酸和镁离子存在的时候合成与模板性质相同的RNA
核心酶(α2ββ ’) 全酶 σ亚基 二、转录 P455页 • 以DNA为模板,按碱基配对原则把DNA的序列信息抄录给RNA,在RNA聚合酶的参与下合成RNA链。 • RNA聚合酶 • 无3´-5’外切酶活性,不需引物;需DNA模板,以NTP(ATP、GTP、CTP、UTP)为底物, RNA链生长方向:5’→3’ • 转录过程: 模板识别、转录的起始、转录的延伸、转录终止
启动子指RNA聚合酶识别、结合和开始转录的一段DNA序列.启动子指RNA聚合酶识别、结合和开始转录的一段DNA序列. RNA聚合酶起始转录需要的辅助因子为转录因子。P459 • 模板识别:RNA聚合酶在σ亚基引导下结合到启动子上 • 双链DNA解开 • 转录的起始:在模板上合成最初RNA(2-9bp) • 转录的延伸:核心酶向前移动,RNA链延长 • 转录终止: 聚合酶达到转录终点 • RNA和聚合酶从DNA上脱落 提供转录停止信号的DNA序列称为终止子,协助RNA聚合酶识别终止信号的辅助因子为终止因子.P464
三、RNA转录后的加工 472页 1.原核生物中RNA的加工 2.真核生物中RNA加工
1.原核生物中RNA的加工 (1)rRNA前体的加工: • 甲基化修饰 • 核酸内切酶消化 • 核酸外切酶消化 (2)tRNA前体的加工: • 核酸内切酶在tRNA的两端切开 • 核酸外切酶从3’端逐个切除附加序列,进行修剪 • 在tRNA的3’端加上-CCAOH • 核苷的修饰 (3)mRNA前体的加工: 大多不需加工,一经转录即可翻译,但也有少数需将多顺反子加工成较小单位。
原核生物rRNA前体的加工 甲基化修饰 RNaseⅢ、RNase E 切割 外切酶切割
核酸内切酶在tRNA的两端切开 核酸外切酶从3’端逐个切除附加序列,进行修剪 ACC 核苷酰转移酶在tRNA的3’端加上-CCAOH 核苷的修饰
2.真核生物中RNA的加工 • rRNA前体的加工: • tRNA前体的加工: • mRNA前体(hnRNA)的加工 • 5’ 端形成特殊的帽子结构 • 在3’ 端切断并加上一个poly(A)的尾巴; • RNA的拼接、编辑和再编码 • mRNA内部甲基化
第三节 蛋白质的生物合成 • 一、遗传密码 • 二、蛋白质合成的分子基础 • 三、翻译的步骤 • 四、蛋白质的运输及翻译后修饰
一、遗传密码 通用遗传密码表 1.遗传密码 • 编码氨基酸的核苷酸序列。反映的是核酸中的核苷酸残基序列与蛋白质中的氨基酸残基序列之间的对应关系。 • 基本单位为三联体密码子
P 513页 2.遗传密码的基本特性 • 密码的简并性 • 密码的变偶性 • 密码子通用性和变异性 • 密码的无标点性、无重叠性 • 密码具有防错系统
P512 密码的简并性 • 同一种氨基酸有两个或者更多的密码子的现象称为密码子的简并性。 • 对应于同一种氨基酸的不同密码子称为同义密码。 • 意义
密码的变偶性 P512 • mRNA 上 密码子专一性取决于前两位碱基,且与tRNA 上 反密码子配对是严格的,第三位碱基可有一定的变动,此现象称。。。 GUA UAA
P513 密码子通用性和变异性 • 密码子的通用性是指各种低等和高等动物,包括病毒细菌和真核生物,公用一套遗传密码。 • 变异性:线粒体DNA以及一些生物中的编码违背了遗传密码的通用性
二、蛋白质合成的分子基础 • mRNA是蛋白质合成的模板 • tRNA转运活化的氨基酸至mRNA模板上 • 核糖体是蛋白质合成的工厂
Phe Phe tRNA 5‘ 3‘ 氨基酸接受臂 1 2 3 5‘ 3‘ 3 2 1 2 tRNA转运活化的氨基酸至mRNA模板上 • 氨基酸接受臂:接受活化氨基酸 • 二氢尿嘧啶环:与氨酰tRNA合成酶的结合有关 • 反密码环:反密码子可以识别信使RNA中的密码子 • 额外环:是分类的指标 • TψC 环:与核糖体的结合有关 书写:
3 核糖体是蛋白质合成的工厂 • 核糖体由大、小两个亚基组成(蛋白质和rRNA) 原核生物:30S 50S 真核生物:40S 60S • 有容纳mRNA的通道 • A位(氨酰基位点)、P位(肽酰基位点) • 能结合起始、延长及终止因子等。 • 有转肽酶活性,催化肽键形成
多核糖体 • 在一条mRNA链上常结合有多个核糖体,呈串珠状排列,同时进行多肽链的合成。
三、翻译的步骤 1.氨酰-tRNA的形成 2.在核糖体上合成蛋白质 mRNA上的阅读方向:从mRNA的5’端→3’端进行。 肽链延伸的方向:从N端→C端
+ 1. 氨酰-tRNA的形成 • 氨酰tRNA合成酶 • 氨基酸 + ATP + tRNA → 氨酰-tRNA + AMP + PPi • 氨基酸 + ATP → 氨酰-AMP + PPi • 氨酰-AMP + tRNA → 氨酰- tRNA + AMP
2. 在核糖体上合成蛋白质 • 起始:起始复合物的形成,在起始密码处开始。 • 延长:进位、转肽、移位 • 终止:到终止密码释放肽链。 全程总结
原核生物翻译起始复合物形成 • IF-1、IF-2、IF-3(initiation factor,IF) • IF-1:促进小亚基与mRNA结合,结合在A位点,阻止氨酰-tRNA进入 • IF-2:促进小亚基与fMet-tRNA结合 • IF-3:促进30S与50S分开,促进小亚基结合mRNA • 过程 • 核蛋白体大小亚基分离; • mRNA在小亚基定位结合; • 起始氨酰-tRNA的结合; • 核蛋白体大亚基结合。
核蛋白体大小亚基分离; • mRNA在小亚基定位结合; • 起始氨酰-tRNA的结合; • 核蛋白体大亚基结合。 IF-2 GTP 5' 3' A U G 过程: IF-2 Pi -GTP GDP IF-3 IF-1
真核生物翻译起始复合物形成 需要更多的蛋白因子(eIF)参与
肽链合成延长 • 在核蛋白体上连续性循环式进行,每次循环增加一个氨基酸: • 进位 • 转肽 • 移位 • 延伸过程所需蛋白因子称为延长因子(elongation factor, EF) • 原核生物:EF-Tu: 促进氨基酰-tRNA进入A位 EF-Ts: 负责催化EF-Tu-GTP形成 EF-G: 转位酶活性,促进mRNA-肽酰-tRNA由A位移到P位 • 真核生物:EF-1 、EF-2 肽酰转移酶
a 进位 根据mRNA下一组遗传密码指导,使相应氨酰-tRNA进入核蛋白体A位。 涉及到的延伸因子:EF-Tu、EF-Ts
5' 3' A U G GTP Tu Ts Ts Tu GDP GTP






















