Download

1 / 1
10 likes | 167 Views
www.rwu.edu. Alkon, TINS, 1998. Bryostatin and lactacystin affect PKC activation and Long Term Memory. Christopher J Gagliardi, Alan Kuzirian*, Herman Epstein*, Daniel Alkon † and Christie Parish § Roger Williams University, One Old Ferry Road, Bristol, Rhode Island 02809

E N D
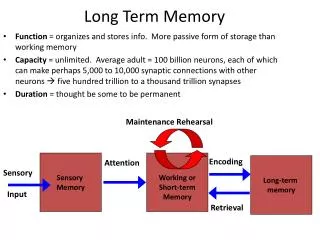
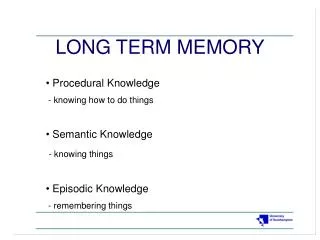
www.rwu.edu Alkon, TINS, 1998 Bryostatin and lactacystin affect PKC activation and Long Term Memory Christopher J Gagliardi, Alan Kuzirian*, Herman Epstein*, Daniel Alkon† and Christie Parish§ Roger Williams University, One Old Ferry Road, Bristol, Rhode Island 02809 Marine Biological Laboratory, 7 MBL street, Woods Hole, Massachusetts 02543 * Blanchette Rockefeller Neurosciences Institute, 9610 Medical Center Drive, Rockville, Maryland 20850 † Nudibranch mollusk Hermissenda crassicornis Effect of 4 Hours of lactacystin plus Bryostatin Followed by Two Training Events Abstract It has recently been shown that protein synthesis plays an important role in the acquisition of sensory learning and the retention of memory. It has also been demonstrated that particular stages of protein synthesis can be manipulated by altering specific components of an established calcium-activated pathway. In this Ca2+ activated pathway, protein kinase C (PKC) is known to function as a key regulator through a direct relationship between PKC activation and the acquisition and retention of memory. bryostatin, a macrocyclic lactone, has been shown to be a potent activator of PKC in sub-molar concentrations. Pavlovian conditioning of the nudibranch mollusk, Hermissendacrassicornis, generated by pairing two natural stimuli (light and agitation) is known to produce an unconditioned response when animals are tested with light alone. Hermissenda have further demonstrated both biochemically and behaviorally, that multiple treatments with bryostatin (0.25ng/mL) administered over several days can induce long-term memory (LTM) from training regimes that typically produce only short-term memory (STM) in untreated animals. Furthermore, to test for the possible relationship between elevated levels of PKC and increased memory retention, we inhibited the ubiquitin-mediated degradation of PKC using the ubiquitin lipase inhibitor lactacystin. Recent results indicate that bryostatin administration, when coupled with lactacystin, increased memory retention by a factor of four compared to animals administered bryostatin (0.25ng/mL) alone. And finally, to demonstrate that protein synthesis was needed for the retention of memory, anisomycin, a protein synthesis inhibitor was used to block mRNA translation. It was found that memory retention dropped significantly when animals were given anisomycin (1ug/mL) immediately following associative training and bryostatin (0.25ng/mL) administration. This present work may have important implications for studies associated with memory acquisition and retention in Alzheimer's patients as the Ca2+ activated, PKC pathway operating in Hermissenda is equivalent in humans. Introduction The focus of our research has been on understanding the process of memory and memory retention. The organism we used to carry out our studies is a small shell-less mollusk called a Hermissenda Crassicornis. Hermissenda are bio-chemically and bio-physically identical and structurally similar to larger vertebrate systems, making them ideal organisms for the study neurological disorders such as Alzheimer’s disease. In fact, Hermissenda possess an identical calcium pathway that is interrupted in Alzheimer’s patients. In this pathway, we focus on protein kinace C (PKC) as a major modulator. There exists a direct relationship between PKC activation and the acquisition and retention of memory. Our Hypothesis is that if the necessary proteins are produced through drug induced PKC activation and are present in subsequent days, the organisms should exhibit long term memory more readily than untreated organisms. The drug we used to test the activation of PKC is Bryostatin (bryo), a cancer drug in stage one trials that is known to activate PKC in small amounts. Bryo increases the translocation of PKC leading to increased levels of calexcitin (CE). We use the immunocytochemical intensity levels of CE as a measure of both PKC and CE. Results show that the short-term effects of repeated bryo exposure down regulates the amount of CE present in the cell. We hypothesized that this down regulation of PKC was a result of the ubiquitin pathway. To test the demonstrated effects of the ubiquitin mediated degradation of PKC, we used the ubiquitin lipace inhibitor lactacystine to inhibit PKC degradation. To test our working bio-chemical pathway, demonstrate that protein synthesis is needed for the retention of memory and to prove that PKC and CE are a part of this pathway; we used an mRNA translation inhibitor called Anisomycin (ANI) to inhibit protein synthesis. Established Ca2+ Activated Pathway Time dependant addition of mRNA translation inhibitor Anisomycin (1ug/mL) Effects of Bryostatin (0.25ng/L) on Memory Retention A.M. Kuzirian, H.T. Epstein H H CHO Br CHO Br CH3 Methods and Materials We used classic Pavlovian conditioning to train our Hermissenda using two natural stimuli paired to cause a conditioned response. Hermissenda are naturally photopositive, and move towards light. Hermissenda are also inter-tidal organisms, when exposed to a stimulus that mimics the ebb and flow of the tide, like a shaking, Hermissenda contract as to secure them self to a substrate. With these two stimuli paired together, the Hermissenda are trained to contract when exposed to light. Each exposure to light paired with a shaking is called a training event (TE). We are able to determine how well an animal remembers training events by exposing the animals to light only and than measuring there body length change by direct measurement. If an animal contracts, it is said to display recall (retention of memory); however, if it does not move, or if it elongates, it is said to display no recall (no retention of memory). Drugs in our experiments are administered by immersion. The drugs are mixed with seawater and than injected into the lanes of Plexiglas trays fixed with openings for syringes. These trays are used for both training and testing the animals. CH3 References 1. Alkon, D. L., Epstein, H. T., Kuzirian, A. M.,Bennett, C.M., Nelson, T. J. 2005. PNAS 102:16432-16437. 2. Etcheberrigaray, R., Tan, M., Dewachter, I., Kuiperi, C., Van der Auwera, I., Wera, S., Qiao, L., Bank, B., Nelson,T.J., Kozikowski, A.P., et al. 2004. PNAS 101:11141-11146. 3. Alkon, D. L., Nelson, T. J., Zhao, W., Cavallaro, S. 1998. TINS 21:529-537. 4. Olds, J.L., Golski, S., McPhie, D. L., Olton,D., Mishkin,M., Alkon, D. L. 1990. J Neurosci 10:3707-3713 5. Muzzio et al., 2001. J Neurophysiol. 85:1639-1647. 6. Mc Phie, D. L., Matzel,L. D., Olds, J. L., Lester, D. S., Kuzirian, A.M. & Alkon, D. L. 1993 J Neurochem. 60:646-651. 7. Bank, B., DeWeer,A., Kuzirian, A. M., Rasmussen, H., Alkon, D. L. 1988. PNAS 85:1988-1992. 8. Scharenberg et al., 1991. PNAS 88:6637-6641. 9. Farley, J., Schuman, E. 1991. PNAS 88:2016-2020. 10. Wender et al., 1998. PNAS 95:6624-6629 11. Crow, T. 2004. Learn Mem 11:31-39. 12. Ramirez, R. R., Gandhi, C.C., Muzziio,I. A., Matzel, L. D. 1998. Learn Mem 4:462-477. Cl Results/Discussion Our working bio-chemical memory pathway begins with the postsynaptic influx of Ca+2, which when coupled with co-factors archidonic acid and inositol triphosphate, induce the autophosphorylation and subsequent translocation of PKC to the cell membrane. The autophosphorylation of PKC causes the phosphorylation of calexcitin (CE) and its subsequent translocation to the cell membrane. CE’s translocation to the cell membrane inhibits potassium channels, which when coupled with internal calcium release, leads to a long lasting depolarization, increasing the resting potential of the cell. This long lasting depolarization is analogous to long-term potentiatian (LTP) in vertebrate systems. Behavioral tests show sub-optimal training normally producing short-term memory (STM) in untreated animals can be induced into long-term memory (LTM) when organisms are treated with bryostatin. Organisms given 6 TE’s with no bryo demonstrae no recall, but with 9TE’s demonstrate significant recall. However, 6 TE’s given with bryo showed an extended memory equal to untreated animals given 9 TE’s. Tests given with the ubiquitin lipase inhibitor lactacystin and bryo paired together have shown far greater memory retention than tests run with bryo alone, further suggesting that PKC plays a major role in the acquisition and retention of memory. Tests given with the mRNA translation inhibitor anisomycin (ANI) and bryo demonstrated recall equal to that of untreated animals, compared to animals given bryo alone with no ANI, which demonstrated significant recall. This adds significant support to our current working model. NH2 OH l OH Acknowledgments Financial support for travel form RWU A special thank you to Alan Kuzirian and Herman Epstein for being terrific mentors,and brilliant scientists Janice Simmons and Nancy Roderic for making my experience at MBL one I will never forget Roxanna Smolowitz for letting me us use her office and microscope for hours on end All of the MRC staff and interns Dr. Cliff Timpson for helping me compile this poster The Marine Biological Laboratory summer internship for making this poster possible