Download

1 / 23
410 likes | 2.12k Views
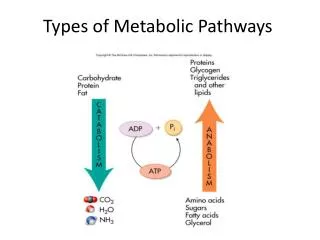
Chapter 23 Metabolic Pathways for Carbohydrates. 23.7 Glycogen Metabolism 23.8 Gluconeogenesis: Glucose Synthesis. Glycogenesis. Glycogenesis : Stores glucose by converting glucose to glycogen. Operates when high levels of glucose-6-phosphate are formed in the first reaction of glycolysis.

E N D
Chapter 23 Metabolic Pathways for Carbohydrates 23.7 Glycogen Metabolism 23.8 Gluconeogenesis: Glucose Synthesis
Glycogenesis Glycogenesis: • Stores glucose by converting glucose to glycogen. • Operates when high levels of glucose-6-phosphate are formed in the first reaction of glycolysis. • Does not operate when energy stores (glycogen) are full, which means that additional glucose is converted to body fat.
Formation of Glucose-6-Phosphate • Glucose is converted to glucose-6-phosphate using ATP. Glucose-6-phosphate
Formation of Glucose-1-Phosphate • Glucose-6-phosphate is converted to glucose-1-phosphate. Glucose-6-phosphate Glucose-1-phosphate
UDP-Glucose • UTP activates glucose-1-phosphate to form UDP-glucose and pyrophosphate (PPi). UDP-glucose
Glycogenesis: Glycogen • The glucose in UDP-glucose adds to glycogen. UDP-Glucose + glycogen glycogen-glucose + UDP • The UDP reacts with ATP to regenerate UTP. UDP + ATP UTP + ADP
Glycogenolysis • Glycogenolysis is the break down of glycogen to glucose.
Glycogenolysis Glycogenolysis: • Is activated by glucagon (low blood glucose). • Bonds glucose to phosphate to form glucose-1-phosphate. Glycogen-glucose + Pi Glycogen + glucose-1-phosphate
Isomerization of Glucose-1-phosphate • The glucose-1-phosphate isomerizes to glucose-6-phosphate, which enters glycolysis for energy production.
Glucose-6-phosphate Glucose-6-phosphate: • Is not utilized by brain and skeletal muscle because they lack glucose-6-phosphatase. • Hydrolyzes to glucose in the liver and kidney, where glucose-6-phosphatase is available providing free glucose for the brain and skeletal muscle.
Utilization of Glucose Glucose: • Is the primary energy source for the brain, skeletal muscle, and red blood cells. • Deficiency can impair the brain and nervous system.
Gluconeogenesis: Glucose Synthesis Gluconeogenesis is: • The synthesis of glucose from carbon atoms of noncarbohydrate compounds. • Required when glycogen stores are depleted.
Gluconeogenesis: Glucose Synthesis • Carbon atoms for gluconeogenesis from lactate, some amino acids, and glycerol are converted to pyruvate or other intermediates. • Seven reactions are the reverse of glycolysis and use the same enzymes. • Three reactions are not reversible. Reaction 1 Hexokinase Reaction 3 Phosphofructokinase Reaction 10 Pyruvate kinase
Gluconeogenesis: Pyruvate to Phosphoenolpyruvate • Pyruvate adds a carbon to form oxaloacetate by two reactions that replace the reverse of reaction 10 of glycolysis. • Then a carbon is removed and a phosphate added to form phosphoenolpyruvate.
Phosphoenolpyruvate to Fructose-1,6-bisphosphate • Phosphoenolpyruvate is converted to fructose-1,6-bisphosphate using the same enzymes in glycolysis.
Glucose Formation • A loss of a phosphate from fructose-1,6-bisphosphate forms fructose-6-phosphate and Pi. • A reversible reaction converts fructose-6-phosphate to glucose-6-phosphate. • The removal of phosphate from glucose-6-phosphate forms glucose.
Cori Cycle • When anaerobic conditions occur in active muscle, glycolysis produces lactate. • The lactate moves through the blood stream to the liver, where it is oxidized back to pyruvate. • Gluconeogenesis converts pyruvate to glucose, which is carried back to the muscles. • The Cori cycle is the flow of lactate and glucose between the muscles and the liver.
Regulation of Glycolysis and Gluconeogenesis • High glucose levels and insulin promote glycolysis. • Low glucose levels and glucagon promote gluconeogenesis.
Ethanol • Ethanol is not a carbohydrate, nor is it a precursor for the biosynthesis of carbohydrates. • However, ethanol can replace sizable amounts of carbohydrates as an energy source when large amounts are ingested. • It is present in the blood of most humans, being produced by intestinal flora. • People ingest ethanol in variable amounts in beverages and fermented fruits. • Ethanol is metabolized in the liver to acetate and adds to the caloric content of the diet. • Ethanol has an energy equivalent of 7 kcal/g. • 100 mL of table wine has ethanol corresponding to about 72 kcal. • A “jigger” of whiskey furnishes approximately 120 kcal.
Ethanol continue: • When ethanol is metabolized in the liver, alcohol dehydrogenase oxidizes it first to acetaldehyde. • CH3CH2OH + NAD+→ CH3CHO + NADH + H+ • The acetaldehyde is oxidized further to acetate. • CH3CHO + NAD+ + H2O → CH3COO- + NADH + H+ • A small fraction of the alcohol may be oxidized by other systems: • Cytochrome P450 oxidase (also involved in detoxification of many drugs); • Catalase • The acetate produced from ethanol largely escapes from the liver and is converted to acetyl CoA and then to carbon dioxide by the way of the Krebs cycle. • The acetyl that stays in the liver may act as a precursor for lipid biosynthesis. • A significant consequence of metabolism of ethanol in the liver is the twofold to threefold increase in the NADH/NAD+ ratio. • With higher concentrations of blood alcohol, the concentration of NADH remains high, and the availability of NAD+ drops and limits both the further oxidation of ethanol and the normal functioning of other metabolic pathways, such as gluconeogenesis.
“Fatty liver” • Chronic consumption of significant amounts of alcohol may lead to a “fatty liver”, in which the excess of triacylglyceride is deposited. • This is caused by several contributing factors: • Reduced triacylglyceride secretion from the liver • Reduced rates of fatty acid oxidation • Increased rates of lipid biosynthesis • These processes are associated with the increased acetyl CoA and NADH/NAD+ ratio in the liver that results from ethanol oxidation.