Download
1 / 81
810 likes | 986 Views
Galvenais audu sader ī bas komplekss. Major Histocompatibility Complex (MHC). Lekcijas plāns. Galvenais audu saderības komplekss (MHC) MHC kompleksa atklāšana un tā loma imūnatbildē MHC organizācija genomā MHC molekulu struktūra Peptīdu saistīšanās pie MHC molekulām

E N D
Galvenais audu saderības komplekss Major Histocompatibility Complex (MHC)
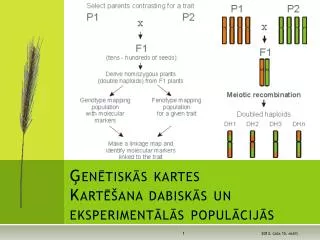
Lekcijas plāns • Galvenais audu saderības komplekss (MHC) • MHC kompleksa atklāšana un tā loma imūnatbildē • MHC organizācija genomā • MHC molekulu struktūra • Peptīdu saistīšanās pie MHC molekulām • MHC molekulu producēšana Antigēna apstrāde un prezentēšana T limfocītiem • To antigēnu īpašības, kurus atpazīst T limfocīti • Antigēnprezentējošās šūnas • Antigēna apstrāde • Lipīdu antigēnu prezentēšana Ilona Mandrikailona@biomed.lu.lv
Kā tika atklāts Galvenais audu saderības komplekss? MHC molekulas tika atklātas pētot audu atgrūšanu peļu līnijās un trušos. • 1916.g. Pārstādot audzējus starp dažādām peļu līnijām • novēroja, ka audzēji tiek pieņemti vienas līnijas ietvaros, bet • atgrūsti dažādu peļu līniju ietvaros. • 1927.g. Secināts, ka pārstādot audus, tie netiek atgrūsti, • ja donors un recipients ir identiski dvīņi. Audu saderība ir atkarīga no ģēnētikas
Kā tika atklāts Galvenais audu saderības komplekss? George Snell un viņa kolēģi (1930-1940) Transplanti vienas peļu līnijas Ietvaros tiek pieņemti. Transplanti starp dažādām peļu līnijām tiek atgrūsti. Transplanta atpazīšana par savu vai svešu ir iedzimta īpašība. Sistemātiski krustoja peļu līnijas, kuras bija ģenētiski identiskas, izņemot vienu ģenētisko rajonu (congenic strains).
Peļu audu saderības lokuss tika sasaistīts ar gēnu 17.hromosomā, kas kodē asins grupas antigēnu (antigēnu II). Šo gēnu rajonu nosauca par Histocompatibility- 2 vai vienkārši H-2. Ģenētisko reģiona daļu, kas kontrolēja audu atgrūšanu un sastāvēja no vairākiem saistītiem gēniem nosauca par Galveno audu saderības kompleksu - Major Histocompatibility Complex (MHC).
Galvenais Audu Saderības Komplekss (Major Histocompatibility Complex) Genoma rajons, kas kodē vairākus gēnus (tāpēc Komplekss), kuri spēlē svarīgu lomu (tāpēc Galvenais) audu translantācijā (tāpēc Audu Saderības).
Turpmākajos 20 gados vienīgā zināmā loma MHC kompleksam bija audu atgrūšana. Transplantācija nav gluži dabīgs fenomens. Kāpēc tad MHC gēni tiek saglabāti evolūcijas gaitā? 1960 - 1970 gados tika atklāts, ka MHC gēniem ir fundamentāla loma visās imūnajās atbildēs pret proteīnu antigēniem.
Cilvēka MHC molekulas = Cilvēka Leikocītu Antigēni Human Leukocyte Antigens HLA = H-2 pelēs • JeanDausset pirmais parādīja, ka indivīdiem, kuri atgrūda nieri, parādījās cirkulējošās antivielas, reaģējošas ar donora limfocītu antigēniem. Šie antigēni (alloantigēni) ir polimorfo gēnu produkti, kuri spēj atšķirt savu no sveša. Pirmie trīs gēni tika nosaukti HLA-A, HLA-B, HLA-C. • BarujBenacerrafatklāja, ka MHC gēni kontrolē • organisma spēju reaģēt uz antigēniem.
Nobela prēmija Fizioloģijā vai Medicīnā 1980. gadā “par viņu atklājumiem attiecībā uz ģenētiski noteiktām struktūrām uz šūnu virsmas, kas regulē imunoloģiskās reakcijas" George D. Snell Jean Dausset Baruj Benacerraf Francija ASV ASV
MHC organizācija genomā
Class II Class III Class I - 20 gēni - 30 gēni - 15 gēni MHC organizācija genomā Cilvēkam MHC atrodas 6.hromosomas īsajā plecā un aptver 3 600 kb (0,1% no cilvēka genoma). Antigēna (peptīda) prezentācija Komplementa sistēmas proteīni Antigēna (peptīda) prezentācija b2-mikroglobulīnu kodē gēns, kas atrodas 15.hromosomā
Class II Class III Class I Klasiskie MHC gēni Class I Class II Kodē gēnus, kas kontrolē visas specifiskās imunatbildes. MHC rajons Kodē arī gēnus, kas ietekmē augšanu, attīstību, vairošanos, smaržas, ožu.
HLA-A HLA-C HLA-B TAP 1 un TAP 2 – transporta proteīnu gēni (nozīmīgi antigēnu pārstrādē) TAP - transporter associated with antigen processing
MHC ir poligēns, t.i. satur vairākus dažādus MHC I un II klases gēnus. MHC ir ar gēniem blīvākais rajons cilvēka genomā.
HLA alēļu skaits (2010.gads) MHC irpolimorfs, t.i.populācijā gēnam ir vairāki varianti. MHC gēni ir vieni no vispolimorfiskākajiem gēniem visā genomā. http://hla.alleles.org/
MHC gēni ir kodominanti ekspresēti katrā indivīdā. Katrā indivīdā vienādā pakāpē tiek ekspresētas mātes un tēva alēles. Tas ļauj palielināt MHC molekulu skaitu nepieciešamo peptīdu piesaistei un T-šūnu prezentācijai. Katrs indivīds ekspresē 6 veidu MHC I klases molekulas uz katras šūnas un 6 vai 8 MHC II klases molekulas.
Šobrīd MHC gēnu lokuss ir viena no visvairāk pētītām ģenētiskām sistēmām. Ietekmē • Infekcijas slimību rezistenci • Autoimunitāti • Audu saderību transplantācijā • Spontānos abortus
Izmaiņas MHC lokusā ir saistītas ar dažāda veida slimībām (>100). ►Monogēnās slimības (hemohromatoze saistīta ar MHCI) ► Infekcijas (HIV-AIDS) ►Autoimūnās slimības (psoriāze, astma, reimatoīdais artrīts) ►Vēzis(Hodžkina limfoma) ►Jūtība pret zālēm
Autoimūnās slimības Vēzis Jūtība pret zālēm Monogēnās slimības Infekcijas
MHC molekulu funkcijas ▶Svarīgas molekulas antigēna atpazīšanā un apstrādē. Inducē un stimulē imūnatbildi. I klases MHC molekulas ▶ Piesaista citoplazmā degradētos antigēnus (vīrusu proteīnus) ▶ Prezentē peptīdus CD8+citotoksiskajiem T- limfocītiem II klases MHC molekulas ▶ Piesaistaendocītiskajos pūslīšos degradētus peptīdus ▶ Prezentē peptīdus CD4+ T limfocītiem palīgiem (T helper)
MHC I un II klases molekulu struktūra
Kopīgas īpašības visām I un II klases MHC molekulām • Transmembrānas glikoproteīni • Ekspresētas uz šūnu virsmas • Ļoti polimorfas • Iesaistītas imunoregulācijā • Katram proteīnam izšķir 4 rajonus - Variablais peptīda-saistīšanas rajons - Konstantais rajons - Transmembrānas rajons - Citoplazmiskais rajons
MHC I molekulas struktūra 2 polipeptīdu ķēdes: ▪ Garā a-ķēde (45kDa) ▪ Īsā ķēde (12kDa) – b2-mikroglobulīns Nekovalenti saistītais b2-mikroglobulīns palīdz stabilizēt MHC molekulas konformāciju. ► Citoplazmiskais rajons sastāv no 30 bāziskām aminoskābēm un satur fosforilēšanas saitus.
MHC I molekulas struktūra ► Transmembrānas rajons – 25 hidrofobās aminoskābes. ►Ļoti konservatīvs a3 domēns pie kura saistās T-šūnu CD8 molekula ►Ļoti polimorfs peptīda saistīšanas rajons, kuru veido abi a1 una2 domēni
MHC II molekulas struktūra Divas aptuveni vienāda garuma polipetīdu ķēdesa unb. • -ķēde ir 32-34 kD b- ķēde ir 29-32 kD ►Īss citoplazmiskais rajons, kas satur fosforilēšanas saitus. ►Transmembrānas rajons – 25 hidrofobās aminoskābes katrā a un b ķēdē.
MHC II molekulas struktūra ►Ļoti konservatīvs b2domēns pie kura saistās T-šūnu CD4 molekula ►Ļoti polimorfs peptīda saistīšanas rajons, kuru veido abi a1 unb1 domēni
Peptīda piesaiste pie MHC molekulas ir fundamentāli atšķirīga no antigēna piesaistes pie B un T limfocītu antigēnu receptoriem • MHC molekulas uzrāda plašu specifiskumu peptīdu piesaistē. Katra MHC molekula var piesaistīt daudz dažādus peptīdus. • MHC molekulas peptīda saistīšanas vietā vienlaicīgi var piesaistīt tikai VIENU peptīdu. • Peptīdiem, kurus saista MHC molekulas ir kopīgas strukturālās īpašības, kuras veicina šo piesaisti
☞MHC I saista 8-11 aminoskābjugarus peptīdus ☞ MHC II saista 10-30 aminoskābju garus peptīdus (optimāli 12-16) ☞ Peptīdi saistās izstieptā konformācijā
Peptīdi saistās pie MHC molekulām caur strukturāli līdzīgām enkura aminoskābēm Katrs peptīds satur 1 vai 2 enkura aminoskābes. Enkura aminoskābes (zaļas) nav identiskas, bet ir līdzīgas: F un Y ir aromātiskas aminoskābes V, L un I ir lielas hidrofobas aminoskābes Ir attēloti peptīdi eluēti no divām dažādām I klases MHC molekulām.
Peptīdu piesaiste pie MHC molekulām • Pie MHC molekulas piesaistītais antigēna peptīds disociē no MHC molekulas ļoti lēni. Kinētikas disociācijas konstantes variē no stundām līdz pat vairākām dienām. • Cilvēka MHC molekulas neatšķir svešus peptīdus no saviem peptīdiem. Kāpēc tad mēs parasti neattīstam imūnatbildi pret saviem proteīniem?
Pilnībā nokomplektētas MHC molekulas ir heterotrimēri MHC I MHC II a- ķēde a- ķēde b- ķēde b2-mikroglobulīns piesaistīts antigēna peptīds piesaistīts antigēna peptīds MHC molekulas bez peptīda ir nestabilas a-ķēdes un b2-mikroglobulīna mijiedarbība pastiprina peptīda antigēna piesaisti Peptīdaantigēna piesaiste stabilizē a-ķēdes un b2-mikroglobulīna mijiedarbību
T-šūnu antigēna receptors atpazīst aminoskābes abos Antigēna peptīdā MHC molekulā Antigēna atpazīšanas specifiskums MHC restrikcija
Self MHC restriction Katra indivīda T-šūnas atpazīst svešus antigēnus TIKAI TAD, ja šie peptīdi ir piesaistīti un izstādīti uz paša indivīda MHC molekulām. Peter Doherty Rolf Zinkernagel 1973-1975.g. atklājumi Nobeļa prēmija 1996. gadā
MHC molekulu ekspresija MHC izplatība ir saistīta ar CD4+ un CD8+ T-šūnu funkcijām. ▶ CD8+ T-limfocīti nogalina šūnas, kuras ir inficētas ar intracelulāriem mikrobiem vai vīrusiem. Vīrusi var inficēt jebkuru kodolsaturošu šūnu. ▶ CD4+ T-šūnas palīdz makrofāgiem iznīcināt ekstracelulāros mikrobus un aktivēt B limfocītus antivielu producēšanai.
MHC molekulu ekspresija MHC I klases molekulas: Ekspresētas uz visām kodolsaturošām šūnām. MHC II klases molekulas: Ekspresētas uz antigēnprezentējošo šūnu virsmas. Tiek regulētas ar IFN-g.
Katra kodolu saturoša šūna ir pārklāta ar vismaz 100 000 MHC I molekulām.
MHC I a ķēdi kodē HLA A HLA B HLA C HLA E HLA F HLA G MHC II a ķēdi kodē HLA DPA1 HLA DQA1 HLA DRA b ķēdi kodē HLA DPB1 HLA DQB1 HLA DRB1 HLA DRB3 HLA DRB4 HLA DRB5
Secinājumi: MHC molekulu struktūra ▶MHC I un II klases molekulām ir ▪ dažāda struktūra, ▪ dažāda izplatība šūnu veidos ▪ dažādas funkcijas ▶Peptīdi, kurus saista MHC I un II klases molekulas ir atvasināti no dažādiem šūnu nodalījumiem un ir dažāda garuma ▶ Interferons-g regulē MHC molekulu ekspresiju
Antigēna apstrāde un prezentēšana T limfocītiem
Antigēnprezentējošās šūnas T- šūnu funkcionēšana ir atkarīga no mijiedarbības ar citām šūnām.
Antigēnprezentējošās šūnas (APŠ) • Pārvērš proteīnu antigēnus par peptīdiem = antigēnu apstrāde. • Eksponē peptīda-MHC kompleksu T-šūnu atpazīšanai. • Ekspresēko-stimulātorus, kuru ir svarīgi T šūnu aktivēšanai. Apoptoze
Dažādu šūnu tipi darbojās kā APŠ lai aktivētu naivās un iepriekš diferencētās T-šūnas Naivās CD4+ Naivās CD8+ Diferenciētas CD4+ T-šūnas T-limfocītus (helper)
Profesionālās APŠ Vienīgās šūnas, kuru galvenās funkcijas ir antigēnu saistīšana un prezentēšana. Vienīgās APŠ, kuras ir spējīgas ierosināt T-šūnu atbildi.
Dendrītiskās šūnas < 1% no šūnu populācijas 25% no epidermas (virsādas) virsmas Dendrītiskās šūnas ierosina T-šūnu atbildi
APŠ antigēn-prezentējošās funkcijas tiek pastiprinātas mikrobu produktu ietekmē MHC un ko-stimulātoru ekspresija Uzlabojās antigēna prezentācijas efektivitāte Citokīnu produkcija APŠ Stimulē T šūnu atbildi Hemokīna receptora CCR7 ekspresija Stimulē APŠ migrāciju uz infekcijas vietām
Naivās T šūnas ekspresē CCR7 un migrē hemokīni
Kāpēc dendrītiskās šūnas ir visefektīvākāsT-limfocītaprimārās atbildes ierosināšanā? • DŠ stratēģiski izvietotas vietās, kur parasti mikrobi iekļūst mūsu organismā. • DŠ ekspresē receptorus, kuri spēj piesaistīt mikrobus (mannozes un Toll-like receptori). • DŠ pārsvarā migrē uz T-šūnu zonām limfmezglos, caur kurām cirkulē naivās T šūnas. • Nobriedūšās DŠ ekspresē kostimulātorus, kuri ir nepieciešami naivo T-šūnu aktivēšanai.
APŠ funkcijas Antigēna apstrāde Antigēna prezentēšana T-šūnām