Download
1 / 12
120 likes | 359 Views
Kromatiini modifitseerimine DNA kahjustuste korral. DNA kaheahelalised katked ( DSB – double-strand breaks ) indutseerivad kromatiini modifikatsioone DNA katkete piirkonnas ning annavad ka rakkudele signaali rakutsükli peatamiseks.

E N D
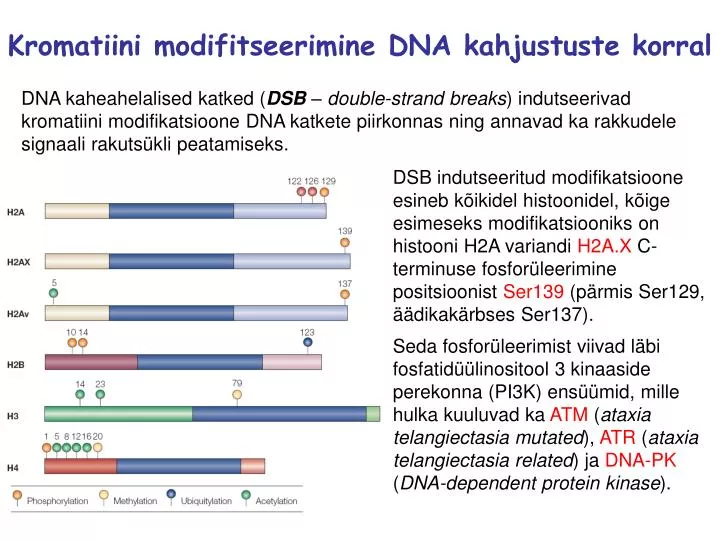
Kromatiini modifitseerimine DNA kahjustuste korral DNA kaheahelalised katked (DSB – double-strand breaks) indutseerivad kromatiini modifikatsioone DNA katkete piirkonnas ning annavad ka rakkudele signaali rakutsükli peatamiseks. DSB indutseeritud modifikatsioone esineb kõikidel histoonidel, kõige esimeseks modifikatsiooniks on histooni H2A variandi H2A.X C-terminuse fosforüleerimine positsioonist Ser139 (pärmis Ser129, äädikakärbses Ser137). Seda fosforüleerimist viivad läbi fosfatidüülinositool 3 kinaaside perekonna (PI3K) ensüümid, mille hulka kuuluvad ka ATM (ataxia telangiectasia mutated), ATR (ataxia telangiectasia related) ja DNA-PK (DNA-dependent protein kinase).
Kromatiini modifitseerimine DNA kahjustuste korral Pärmis teevad sama töö ära kinaasid Tel1 ja Mec1 ning fosforüleeritud histooni H2A võib leida kuni 50 kb kaugusel DNA katkemise kohast (inimese rakkudes leidub fosfo-H2A.X isegi kuni 2 Mb kaugusel DSB-st), kõrgeim H2A fosforüleerituse tase on umbes 3-5 kb kaugusel DSB-st, kuid huvitaval kombel on fosfo-H2A tase vahetult DNA katkestuse läheduses (1-2 kb ulatuses) suhteliselt madal. H2A.X fosforüleerimissaidiga mutantsed pärmi- ja hiirerakud on ioniseerivale kiirgusele (tekitab DSB) tundlikumad kui wt histoonidega rakud. Arvatavasti on H2A.X fosforüleerimine vajalik DNA reparatsiooniks oluliste faktorite stabiilseks seondumiseks kahjustatud DNA alaga. Samas pole H2A.X fosforüleerimine vajalik nende faktorite algseks kohaletoomiseks.
Kromatiini modifitseerimine DNA kahjustuste korral H2A.X fosforüleerimine on ilmselt vajalik histooni atsetüültransferaasi NuA4 ning kromatiini remodelleerivate komplekside INO80 ja SWR1 seondumiseks Kõikide nende komplekside ühisosa on Arp4 valk (actin-related protein 4). NuA4 atsetüleerib histooni H4 N-terminuses nelja esimest lüsiinijääki. Kui kõik 4 lüsiinijääki on muteeritud, või kui NuA4 on inaktiveeritud, siis muutuvad rakud tundlikuks DNA DBS tekitavate kemikaalide suhtes. Ilmselt on NuA4 poolt tehtav H4 atsetüleerimine vajalik kromatiini avamiseks DNA kahjustuse piirkonnas. Samas on leitud, et ka histoonide deatsetülaasi kompleksi Sin3/Rpd3 inaktiveerimine põhjustab häireid DSB parandamises. Histoonide atsetüleerituse tase DNA ahelate katkemiskohas võib olla seotud valikuga, kas vigastus parandatakse NHEJ või HR süsteemiga: HR korral on histoonide atsetüleerituse tase kõrgem.
Kromatiini modifitseerimine DNA kahjustuste korral INO80 ja SWR1 on kromatiini remodelleerivad kompleksid ning sisaldavad subühikuid Rvb1 ja Rvb2, mis on väga sarnased bakteriaalsele RuvB valgule. RuvB aga on vajalik Holliday ühenduste moodustamiseks ja liigutamiseks. See annab põhjust oletada, et INO80 ja SWR1 on seotud kromatiini remodelleerimisega DSB reparatsiooni korral. Seda toetab ka fakt, et kui rakkudes puudub kas funktsionaalne INO80 või SWR1, siis nad on tundlikud DSB tekitavate kemikaalide suhtes. INO80 tulekuks DSB piirkonda on vajalik histooni H2A.X fosforüleerimine: kui inaktiveerida Tel1 ja Mec1 kinaasid või kui rakkudes on mittefosforüleeritav H2A.X mutant, siis INO80 tulek DNA kahjustuste piirkonda on oluliselt häiritud. Milleks kromatiini remodelleerijaid vaja on? INO80 puudumisel toimub DNA otsa protsessing (ssDNA teke) ebaefektiivselt, mis näitab, et remodelleerijaid on vaja eeskätt DNA vabastamiseks nukleosoomidest ning reparatsioonifaktoritele ligipääsu tagamiseks.
Kromatiini modifitseerimine DNA kahjustuste korral Mõned kromatiini remodelleerijad seonduvad DSB piirkonnaga ka fosfo-H2A.X-st sõltumatult. Näiteks RSC kompleks tuleb kohale koos MRX ja Ku kompleksidega, mis on esimesed, kes katkenud DNA otstega seonduvad. Fosfo-H2A.X on vajalik ka kohesiini lokaliseerumiseks DNA katkete piirkonda. Kohesiin on oluline tütarkromatiidide omavaheliseks sidumiseks, mis on eriti oluline homoloogse rekombinatsiooni kaudu DNA katkete parandamisel.
Kromatiini modifitseerimine DNA kahjustuste korral Kui DNA vigastus on parandatud, siis on oluline vabaneda fosfo-H2A.X-st, sest fosfo-H2A.X on rakkudele signaaliks katkenud DNA-st ning selle signaali tulemusena peatatakse rakutsükkel. Fosfo-H2A.X-st vabanemiseks on kaks võimalust: 1. See asendatakse H2A.Z-ga, mis on teine H2A variant ning mida lülitatakse kromatiini SWR1 kompleksi poolt. 2. H2A.X defosforüleeritakse PP2A(C) fosfataasi poolt. Ka histoonide atsetüleeritus eemaldatakse Sin3/Rpd3 HDAC kompleksi poolt, lisaks toimub histooni H4 Ser1 fosforüleerimine CK2 poolt, mis takistab NuA4-l histoon H4 uuesti atsetüleerida.
Teised histoonide modifikatsioonid DNA katketega kaasneb ka palju muid histoonide modifikatsioone. Enamusel juhtudel pole teada, kas tegu on lihtsalt DNA reparatsiooniga kaasnevate modifikatsioonidega, või on neil aktiivne roll DNA reparatsiooni reguleerimisel. On leitud, et rakutsükli G2 faasis püsib histoon H3 lüsiin 56 atsetüleerituna seni, kuni DNA vigastused on parandatud. Lys56 atsetüleeritakse kohe peale histooni H3 sünteesi, kuid seda modifikatsiooni ei eemaldata vahetult peale histooni lülitamist kromatiini, vaid see jääb püsima S faasi lõpuni. AcK56 eemaldatakse alles siis, kui kõik DNA replikatsiooni käigus tekkinud DNA katked on parandatud. Seega on H3 Lys56 deatsetüleerimine märgiks, et rakk on edukalt läbinud S-faasi “kvaliteedikontrolli”. Pole teada, mida see modifikatsioon konkreetselt teeb või millega seondub, kuid on teada, et H3 Lys56 mutandid on DNA katkeid põhjustavate kemikaalide suhtes isegi tundlikumad, kui H2A.X fosforüleerimise mutandid. Ka histooni H3 lüsiin 79 metüleerimisel on seos DNA vigastuste kontrolliga. Lys79 metüleerimine küll ei muutu sõltuvalt DNA kahjustusest, kuid see modifikatsioon on vajalik teiste valkude seondumiseks kromatiiniga. Metüleeritud Lys79 tunnevad ära mõned DNA reparatsiooniks ja S-faasi kontrolliks olulised valgud, näiteks 53BP1.
Teised histoonide modifikatsioonid Sarnaselt H3 K79 metüleerimisega on leitud, et ka histooni H4 lüsiin 20 metüleerimine on seotud DNA reparatsiooniga. Ka sellel juhul on tegu modifikatsiooniga, mis ei ole otseselt indutseeritud DNA katkete poolt, kuid see modifikatsioon on oluline seondumiskoht S-faasi kontrollvalgule Crb2. Arvatavasti on H3 K73 ja H4 K20 metüleerimised olulised kromatiini kõrgemat järku struktuuride kontekstis. Mõlemad lüsiinid on 30-nm fiibri struktuuris raskesti ligipääsetavad, kuna jäävad maskeerituks naabernukleosoomi poolt. DNA katkestused aga tekitavad muudatusi ka 30-nm fiibri struktuuris ning K79 ja K20 muutuvad teistele valkudele kättesaadavaks. Kui rakkudes puudub H3 K79 metüleerimine, siis ei suuda S-faasis toimuvad DNA katked käivitada S-faasi kontrollsüsteemi ning rakutsükkel ei peatu, et DNA vigastusi parandada
Rakutsükli kontrollpunktid Kontrollpunkide aktiveerimine viib kas rakutsükli aeglustumiseni, või selle täieliku peatumiseni. Näiteks kui S-faasis avastatakse DNA katked, siis uusi replikatsiooni origin’e ei aktiveerita, küll aga lastakse DNA süntees lõpetada nendel polümeraasidel, mis on juba alustanud. Kontrollpunktide signaalirada on väga tundlik, näiteks ATM-i ja S-faasi kontrollpunkti aktiveerimiseks piisab vaid mõnest DNA katkest.
Nukleosoomid ja DNA reparatsioon ssDNA kahjustuste korral on täheldatud, et need kromosoomi alad, mis jäävad heterokromatiini piirkonda, parandatakse aeglasemalt kui need, mis on eukromatiinis. Samas on ka leitud, et nukleosoom võib mingil määral kaitsta DNA-d kahjustuste eest. Näiteks UV-st tekitatatud CPD-d paiknevad keskmiselt iga 10,3 bp tagant. Täpselt sama pika vahemaa tagant toimub DNA seondumine nukleosoomile. Üldjuhul tekitab nukleotiidiga toimunud modifikatsioon DNA struktuuris lokaalseid muudatusi ning see võib iseenesest põhjustada problemaatilise nukleotiidi “eksponeerimise” reparatsiooniensüümidele. DNA vigastuse asukoht nukleosoomis võib mõnedel juhtudel mõjutada vea parandamise kiirust. Näiteks need CPD-d, mis asuvad nukleosoomi keskel parandatakse fotolüaasi poolt aeglasemalt, kui need, mis asuvad nukleosoomi äärealadel.






















