Download

1 / 25
250 likes | 704 Views
Cellens biologi og patologi. Kap. 3 i Almen patologi, 2. udgave. Celler . Celler er de mindste, levende, selvstændigt, reproducerende, funktionelle enheder.

E N D
Cellens biologi og patologi Kap. 3 i Almen patologi, 2. udgave
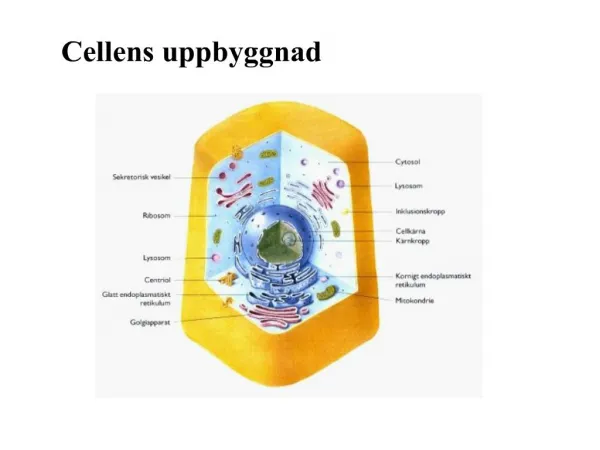
Celler Celler er de mindste, levende, selvstændigt, reproducerende, funktionelle enheder. Cellens cytoplasma har en høj grad af funktionel og strukturel specialisering, idet forskellige typer af membranafgrænsende strukturer, organeller, og ikke-membranafgrænsende strukturer som f.eks. mikrofilamenter varetager bestemte funktioner. Cellens mange funktioner styres af gener i cellens DNA som via messenger RNA og ribosomalt RNA katalysere dannelsen af de mange tusind proteinmolekyler. Forbigående eller varige ændringer i genernes funktion spiller en central rolle ved måske alle sygdomme, såvel erhvervede som arvelige.
Cellekerne - kromatin Kernen er afgrænset af en kernemembran og indeholder de følgende komponenter: • Kromatin – DNA-holdige kernemateriale. Udover DNA, består kromatin af RNA og proteiner. DNA danner lange negative ladede kæder, som er viklet omkring partikler af otte positivt ladede proteinmolekyler (histoner), de såkaldte nukleosomer. Disse fibre samles igen i en spiralstruktur til ca. 30 nm tykke kromatintråde, som ved yderligere snoning danner kromosomer. Der er 2 slags kromatiner: • Eukromatin – er løspakket og er transkriptionelt aktivt • Heterokromatin - er tætpakket og er transkriptionelt inaktivt og findes oftest perifert i kernen umiddelbart op ad kernemembranen samt omkring nucleolus. Det kan inddeles yderligere i 2 slags: • Konstitutivt – altid inaktivt og overvejende repetitivt DNA • Fakultativt – kan være aktivt eller inaktivt.
Cellekerne - kernematrix • Kernens matrix - udgøres af et netværk af proteiner, som danner kernens skelet og bestemmer kernens størrelse og form. Matrixproteinerne deltager i en række af kernens funktioner, herunder replikation og transkription, og fungerer som bindingssted for enzymer og andre proteiner, som deltager i disse processer. Ved neoplasi ændrer disse proteiner og er medvirkende til at tumorcellers kerner morfologisk afviger fra normale cellers. • Nucleolus – består af ribonukleoprotein, det nucleolusassocierede kromatin samt en proteinmatrix. Nukleoler dannes på 5 bestemte kromosomer fordelt på 10 regioner som indeholder såkaldte nukleoleorganisationsregioner, NOR. De fleste er permanent inaktiverede og i humane celler ses der kun 1-2 eller sjældent mere end 4 nukleoler. NOR består af store slynger af ribosomalt DNA og er af centralt betydning for transkriptionen af rRNA som danner ribosomer sammen med protein fra cytoplasmaet. I forbindelse med neoplasi ses øget antal NOR. Disse kan påvises ved sølvfarvning som AgNOR
Kernemembran Kernemembranen består af 2 koncentrisk lejrede enkeltmembraner. Den inderste kan på indersiden være dækket af et elektrontæt lag. Mellemrummet mellem disse to membraner, det perinukleære rum, står i forbindelse med det granulære ER, idet den yderste membran fortsætter som membran i det granulære ER. Kernemembranen indeholder talrige porer, hvor der foregår transporten af molekyler mindre end 60 kD til og fra kernen.
Cytoplasma og organeller Cytoplasmaet udgør den største del af cellen. Det omgiver kernen og afgrænses fra det ekstracellulære rum af cellemembranen. Der forekommer forskellige organeller i cytoplasma: • Endoplasmatiskreticulum • Granulært endoplasmatiskreticulum – består af ribosombeklædte membraner, som omslutter fladeformede sammenhængende cisterner. De frie ribosomer er 25 nm store partikler, som består af rRNA der vha. messenger og tRNA katalyserer proteinsyntese. De katalyserer dannelsen af de mange proteiner, bl.a. enzymer som indgår i opbygning og fornyelse af alle cellens cytoplasma- og kernestrukturer. • Glat endoplasmatiskreticulum – er involveret i metaboliske processer f.eks. lipidmetabolisme, dannelse af steroidhormoner og i leveren i nedbrydning af medikamenta og toksiner.
Cytoplasma og organeller • Golgi-apparat – proteiner som dannes i det ru ER, passeres via overgangsvesikler til Golgi-cisterner på disse modtageflade. Her foregår modifikation af kulhydratdel af molekylet. De membranstrukturer, som afsnøres fra Golgi-cisterner, omfatter forskellige mere eller mindre tæt pakkede vesikler, som enten er sekretvesikler og granula med materiale beregnet til brug uden for cellen eller som cellen selv bruger. • Lysosomer – disse dannes ved modning af Golgi-vesikler. De har et højt indhold af hydrolyserende enzymer, som ved surt pH nedbryder organiske forbindelser. De spiller en vigtig rolle i cellernes protein-, kulhydrat- og lipidomsætning, idet de som fagolysosomer nedbryder endocyteret materiale. Ligeledes er omsætningen i autofagolysosomer af cellens egne bestanddele vigtig. Sur fosfatase og katalase er de bedste kendte lysosommarkører.
Cytoplasma og organeller Mitokondrier – de består af 2 membransystemer, den inderste omslutter mitokondriematrix og danner folder, de såkaldte mitokondriecristae. Den yderste omslutter den inderste membran. Langt hovedparten af cellens oxidative stofskifte foregår i mitokondrierne, hvorved der bl.a. dannes adenosintrifosfat (ATP). • Peroxisomer – navnet skyldes indhold af enzymer, der er involveret i omsætningen af hydrogenperoxid, herunder katalase. Den findes i alle kroppens celler, flest i lever og nyrer. De dannes ved vækst og deling fra allerede eksisterende peroxisomer. Peroxisomer udfører en række enzymatiske reaktioner, som f.eks. nedbrydning af alkohol og fedtsyrer samt syntese af galdesyrer. Zellwegers syndrom er karakteriseret ved total mangel på peroxisomer, der viser sig ved kraniofacialdysmorfi, svær hypotoni og manglende psykomotorisk udvikling og børnene lever sjældent mere end et halvt år.
Lipofuscin De lysosomale nedbrydningsprodukter bruges normalt igen af cellen, men ofte bliver små rester af komplekse, ikke-nedbrydelige lipidforbindelser tilbage som gulligt-brunecytoplasmagranula, der benævnes lipofuscingranula eller alders pigment. Disse består hovedsageligt af membranmateriale fra mitokondrier og cellemembraner. Lipofuscinophobning har næppe betydning for cellernes funktion.
Sekretgranula Sekretgranula dannes ligesom lysosomer. Mens lysosomer forbliver i cellen, passerer sekretgranula fra Golgi-apparatet til celleoverfladen. Transporten ud af cellen kaldes exocytose. Identifikation af granulas indhold foretages som regel ved immunhistokemisk undersøgelse.
Cytosol Cytosol er cytoplasmaet mellem cellens organeller og udgør ca. ½-delen af cellevolumen. Det består af forskellige proteinstrukturer, det såkaldte cytoskelet, samt af ikke strukturerende proteiner som er flydende og derfor muliggør strømning og transport i cytoplasmaet.
Cytoskelet Cytoskeletet er med til at opretholde cellens form og er en integreret del af cellefunktioner som cellebevægelse, endocytose, exocytose og celledeling. Cytoskeletet udgøres af de stabile intermediærfilamenter, og de mere labile aktinfilamenter og mikrotubulier: • Intermediærfilamenter – er 10 nm i diameter, er permanente og har overvejende stabiliserende mekaniske funktioner, dvs. at de ikke hurtigt dannes og nedbrydes som mikrotubuli og aktinfilamenter. • Aktinfilamenter – 6 nm i diameter, findes i alle celler enten alene eller i forbindelse med myosinfilamenter. Dødsstivhed, rigormortis, skyldes at aktin bindes til myosin. I muskelceller danner disse med myosin filamenter kontraktile enheder. I andre celler indgår de i den terminale afsnøring af cellen ved celledeling. De danner også den aksiale struktur i mikrovilli i f.eks. tarmens absorptive celler. • Mikrotubuli – 23 nm i diameter, cytoplasmas mikrotubuli er labile, idet de hurtigt kan vokse i længden ved polymerisering af frit tubulin og tilsvarende hurtigt depolymerisere. Disse fungerer dels som støttefibre, dels som strukturer langs hvilke cytoplasmaets strømninger og stoftransport foregår. De dannes også i forbindelse med celledeling, som udgør cellens mitosespindelfibre, styret af centrioler.
Cellemembran Cellemembranen afgrænser cellens indre miljø og dermed de markante ionkoncentrationsforskelle som findes mellem det ekstracellulære og det intracellulære rum. Som alle biologiske membraner er cellemembranen opbygget af lipider og proteinmolekyler. Lipidmolekylerne er altid arrangeret i to lag, som holdes sammen af non-kovalente bindinger, der dog ikke er stærkere end at der er fri bevægelighed inden for hvert lag i det dobbelte lipidlag. Cellemembranen har herudover en lang række funktioner med hensyn til transport af stoffer ind og ud af cellen, overførelse af informationer, antigen egenskaber og bindinger mellem celler og ekstracellulære strukturer. Lipiddelen af membranen danner en barriere, som er impermeabel for hydrofile og store uladede, polære molekyler samt ionmolekyler, og der findes i cellemembranen nogle transmembrane transport- og kanalproteiner, som er ansvarlige for transporten af disse. Alle cellemembraners overflade er dækket af glykocalyx
Basalmembran Basalmembraner er stabile bindevævsstrukturer, som danner et sammenhængende lag under alle epitelceller undtagen leverceller, og under alle endotelceller undtagen sinusoider i leveren, i knoglemarven og i lymfoide organer. Basalmembraner syntetiseres af de celler, som hviler på dem. Elektromikroskopisk består basalmembraner af en 80-100 nm tyk fortætning af fine filamenter indlejret i en amorf matrix. Denne betegnes basallamina, og på udsiden af denne findes en bredere zone af fibre, som kaldes den retikulærelamina. Basalmembranen består af kollagen type IV, proteoglykaner som heparansulfat samt laminin. Basalmembraner fungerer som støtte for de ovenliggende celler og medvirker også ved regeneration af disse celler, formentlig fordi basalmembraner kan binde vækstfaktorer. Basalmembraner fungerer også som filter for stofpassage. Svigt af denne filterfunktion forklarer, hvorfor fortykkelse af basalmembraner kan ses samtidig med, at disses permeabilitet er øget.
Ekstracellulær matrix Ekstracellulær matrix består af et kompleks af makromolekyler, som dels findes som fibre, dels som en amorf grundsubstans. Den er syntetiseret af cellerne lokalt. Cellulær matrix findes i bindevæv. Fibre i den ekstracellulære matrix omfatter kollagen og elastin.
Kromosomer I menneskets celler er der 46 kromosmer, der danner 23 homologe par, som hver består af maternelt og paternelt kromosom. Hvert kromosom består af 2 ens kromatider, der er forbundet ved centromeret. Herved opstår 2 sæt arme, hvoraf den korte kaldes for p-armen og den lange for q-armen. Hvert arm afsluttes af telomeret.
DNA Et komplet sæt af humant DNA omfatter ca. 2,85 mia. basepar og den normale diploide celle indeholder derfor ca. 5,700 Mbp DNA. Af den totale DNA-mængde findes ca. 20% i generne og heraf udgør de kodende dele (exons) ca. en fjerdedel. Resten af DNA består dels af repetetive DNA-sekvenser (ca. 20%) og junk-DNA (ca. 60%) (ukendt funktion).
Satellitter De 20% af repetetiveDNA-sekvenser findes i centromerer og telomerer og spredt på kromosomerne. Efter deres størrelse kan disse deles i megasatellitter, satellitter, minisatellitter og mikrosatellitter. Satellitter er meget polymorfe, dvs. at der kan findes 10 eller flere alleler af forskellig længde. Usystematiske ændringer i mikrosatellitter fra celle til celle (mikrosatellit-instabilitet, MSI) kan være udtryk for mutationer i bestemte DNA-reparationsgener (mismatchrepair-gener). Da sådanne mutationer kan medføre fejl i replikationen, kan de også medføre mutationer i andre gener bl.a. betydning for udvikling af cancer.
Cellecyklus og celledeling Tiden fra en til den efterfølgende celledeling kaldes cellens cyklus. Cellens cyklus omfatter 4 faser: • Interfasen • G1-fase – længden af denne fase afgør forskellen mellem de labile, stabile og permanente celler. • S-fasen – her foregår DNA-replikation. processen katalyseres af enzymet DNA-polymerase. Denne fase kan vare ca. 7 timer • G2-fasen – her foregår reparation af defekt DNA. Denne fase kan vare 3 timer • M-fasen – her bliver kromosomerne synlige som strenge eller tråde G0-fasen er den, hvor cellerne ikke deler sig. Overgangen mellem de forskellige faser reguleres af såkaldte checkpoint og stimuleres bl.a. af vækstfaktorer.
Cellecyklus og celledeling • Profase – her ses kromosom som to kromatider, forbundet ved centromeret • Metafase • Anafase • Telofase • Cytokinesen – her deles cytoplasma
Celledød Celledød kan foregå på 2 måder: • Apoptose • Nekrose Mens apoptose induceres af såvel fysiologiske som ufysiologiske stimuli, er nekrose udelukkende en følge af ufysiologiske stimuli. Apoptose medvirker under følgende forhold: • Embryogenesen • Opretholdelse af den normale homeostase • At fjerne beskadigede eller slidte celler • Den normale aldringsproces
Mutationer Der sker flere millioner DNA-skader hvert år, de fleste i forbindelse med S-fasen før celledelingen. Næsten alle disse skader repareres af cellens DNA-reparationsenzymer. De fleste af disse skadede celler går til grunde, men enkelte overlever med en permanent ændring i sit DNA, dvs. en mutation. I hvert gen er der beskrevet ca. 100 mutationer, hvoraf de fleste er betydningsløse da de sker i ikke-kodende dele af DNA. Andre kan dog medføre alvorlige skader og sygdomme. Mutationer i mtDNA er hyppigere end i nukleært DNA, da førstenævnte mangler beskyttende histoner og DNA-reparationssystemer.
Mutationer Mutationer kan inddeles på flere måder: • Kromosomantalsmutationer f.eks. abnormaltkromosomtal. • Kromosommutationer f.eks. strukturelle forandringer som brud på kromosomerne bliver repareret ukorrekt. • Genmutationer - f.eks. punktmutationer, frameshift mutation, genamplikation, missense mutation, nonsense mutation • Tab af heterozygositet LOH – LOH i tumorceller er som regel tegn på tab af anti-onkogener. LOH spiller en rolle ved sygdomme retinoblastom og kolerektalcancer.
Mutationer Årsager til mutationer: • Stråling • Vira • Kemiske stoffer • Cytostatika • Mutagene stoffer – fremkalder mutationer • Teratogene stoffer – fremkalder misdannelser hos fostret • Karcinogener – fremkalder maligne tumorer • Nogle arvelige sygdomme, der kan medføre cancersyndromer pga. mutation. I næsten alle disse tilfælde drejer det sig om inaktivering af tumorsuppressorgener.
Påvisning af mutationer Mutationer og deres følger kan påvises med mange forskellige metoder, som f.eks. cytogenetisk undersøgelse (karyotypering), FISH og PCR, og i nogle tilfælde ved molekylærbiologiske teknikker.