Download
1 / 31
430 likes | 1.07k Views
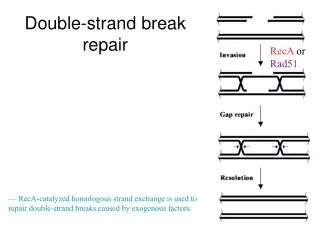
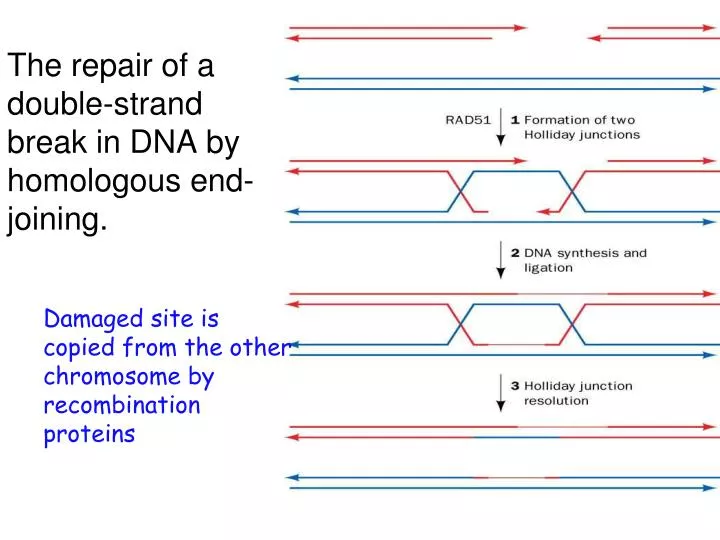
The repair of a double-strand break in DNA by homologous end-joining. Damaged site is copied from the other chromosome by recombination proteins. Double strand repair. Nonhomologous end-joining only in emergency situations Two broken ends of DNA are joined together.

E N D
The repair of a double-strand break in DNA by homologous end-joining. • Damaged site is copied from the other chromosome by recombination proteins
Double strand repair • Nonhomologous end-joining • only in emergency situations • Two broken ends of DNA are joined together. • A couple of nucleotides are cut from both of the strands. • Ligase joins the strands together. • Sometimes an extra nucleotide is added.
Construct design in this paper is more complex, but the idea is the same. Reporter gene is GFP. Since repair is taking place in a eukaryotic system, ds breaks must be repaired, mRNAs synthesized and processed by normal splicing machinery to generate detectable levels of GFP. SceI sites produce HR sites in Construct B (next slide) and NHEJ type breaks in construct A. Constructs are integrated into the chromosomes Sce I site
SD, splice donor; SA, splice acceptor; shaded squares, polyadenylation sites. Supplementary Figure 1. Constructs integrated in the reporter cell lines for detecting NHEJ and HR. A, NHEJ reporter cassette. The construct consists of a GFP gene containing an intron, interrupted by an adenoviral exon (Ad).The adenoviral exon is flanked by SceI recognition sites in inverted orientation for induction of DSBs. Inverted orientation prevents overlap of sticky ends produced by SceI cleavage (see Figure 1C). In the starting construct the GFP gene is inactive. Induction of a DSB by I-SceI followed by NHEJ, transcription and splicing reconstitutes the functional GFP gene.
SceI recognition site …TAGGGATAACAGGGTAAT…ATCCCTATTGTCCC ATT… …ATCCC TATT GTCCCATTA…TAGGGATAACAGGGTAAT Two SceI sites in inverted orientation make incompatible sticky ends.
B, HR reporter cassette. The construct consists of two mutated copies of GFP-Pem1. In the first copy of GFP-Pem1 the first GFP exon carries a deletion of 22 nt and an insertion of two I-SceI recognition sites in inverted orientation. The 22 nt deletion ensures that GFP cannot be reconstituted by an NHEJ event. The second copy of GFP-Pem1 is lacking the ATG (for translation initiation) and the second exon of GFP. Upon induction of DSBs by I-SceI, gene conversion events reconstitute the GFP gene.
The construct consists of two mutated copies of GFP-Pem1. In the first copy of GFP-Pem1 the first GFP exon carries a deletion of 22 nt and an insertion of two I-SceI recognition sites in inverted orientation. The 22 nt deletion ensures that GFP cannot be reconstituted by an NHEJ event. The second copy of GFP-Pem1 is lacking the ATG and the second exon of GFP. Upon induction of DSBs by I-SceI, gene conversion events reconstitute the GFP gene. #1 #2
Fig. 1 SIRT6 stimulates DSB repair. Published by AAAS Z Mao et al. Science 2011;332:1443-1446
Fig. 1 SIRT6 stimulates DSB repair. A) Overexpression of SIRT1, -2, -6, and -7 in human fibroblasts. Immunoblotting with sirtuin-specific antibodies after transfection with a sirtuin-expressing vector or a control vector encoding hypoxanthine-guanine phosphoribosyltransferase (pControl). Z Mao et al. Science 2011;332:1443-1446 Z Mao et al. Science 2011;332:1443-1446 Published by AAAS
Fig. 1 SIRT6 stimulates DSB repair. (B) Effect of sirtuin overexpression on the efficiency of NHEJ and HR, measured as described in (27) and fig. S1. The efficiency of DSB repair was scored in untreated cells (open bars), cells pretreated with 1 mM paraquat for 16 hours(black bars), or cells treated with paraquat and 5 mM nicotinamide for 16 hours (red bars). Error bars indicate SD; n = 8 experiments (control and SIRT6); n = 3 (other sirtuins). P values were calculated by two-tailed Student’s t test.. Z Mao et al. Science 2011;332:1443-1446 Published by AAAS
Fig. 1 SIRT6 stimulates DSB repair. (C) SIRT6 overexpression accelerates the disappearance of γH2AX foci after treatment with 1 mM paraquat for 16 hours. Data represents an average of at least 50 nuclei.
Fig. 1 SIRT6 stimulates DSB repair. (D) Immunoblot showing induction of endogenous SIRT6 protein levels by oxidative stress. Human fibroblasts were treated with paraquat for 16 hours. Z Mao et al. Science 2011;332:1443-1446 Z Mao et al. Science 2011;332:1443-1446 Published by AAAS
Fig. 2 Oxidative stress results in early recruitment of SIRT6 to DNA breaks. Oxidative stress results in early recruitment of SIRT6 to DNA breaks. ChIP analysis showing kinetics of SIRT6 recruitment to Alu sequences after 8 Gy of γ-irradiation (IR) (A) and sequences flanking I-SceI–induced DSB after transfection with I-SceI expression vector (B). Asterisks indicate values significantly different from corresponding zero time points (P < 0.05). Error bars indicate SD; n = 5. Control ChIP with SIRT6−/− cells is shown in fig. S7. IgG, immunoglobulin G. Z Mao et al. Science 2011;332:1443-1446 Published by AAAS
Fig. 3 Deacetylation and mono-ADP-ribosylation activities of SIRT6 are required to stimulate DSB repair. Immunoblot showing that S56Y and R65A mutations abolish the H3K9 deacetylation activity of Sirt6 and appear to exert a dominant-negative effect. (B) In vitro assay showing that S56Y and G60A mutations abolish mono-ADP-ribosylation activity of SIRT6. NAD+, nicotinamide adenine dinucleotide. Z Mao et al. Science 2011;332:1443-1446
Fig. 3 Deacetylation and mono-ADP-ribosylation activities of SIRT6 are required to stimulate DSB repair. (A) Immunoblot showing that S56Y and R65A mutations abolish the H3K9 deacetylation activity of Sirt6 and appear to exert a dominant-negative effect. (B) In vitro assay showing that S56Y and G60A mutations abolish mono-ADP-ribosylation activity of SIRT6. NAD+, nicotinamide adenine dinucleotide. (C) SIRT6 mutants for deacetylation and/or ribosylation activities have reduced ability to stimulate NHEJ and HR. Untreated cells (open bars) or cells treated with paraquat (black bars) were transfected with SIRT6-expressing vectors or pControl. Z Mao et al. Science 2011;332:1443-1446
Fig. 4 SIRT6 interacts with PARP1 and stimulates its poly-ADP-ribosylation activity. Z Mao et al. Science 2011;332:1443-1446 (A) Analysis of mono-ADP-ribosylated proteins in the WT and SIRT6−/− MEFs stressed with paraquat for 16 hours. Cells were transfected with biotinylated NAD, and poly-ADP-ribosylated proteins were cleared away with PAR antibodies. (B) Immunoblotting of the extracts in (A) with PARP1 antibodies indicated that the 120-kD band is Parp1. (C) SIRT6 interacts with PARP1. Human fibroblasts were treated with 1 mM paraquat. Cell lysates were immunoprecipitated with SIRT6 antibodies in the presence of ethidium bromide (EtBr) followed by Western blotting with PARP1 antibodies.
(E) PARP1 lacking the catalytic domain is mono-ADP-ribosylated by SIRT6 in vitro, whereas K521A is not. (F) In vitro assay of PARP1 poly-ADP-ribosylation activity showing that PARP1 is stimulated only by SIRT6 mono-ADP-ribosylation activity. (G) Stimulation of NHEJ and HR by SIRT6 is abolished by PARP1 inhibitors 3-ABA or PJ34. (D) PARP1 K521 is essential for activation of NHEJ by SIRT6. An NHEJ assay was performed in PARP1−/− MEFs containing an integrated NHEJ reporter. Cells were transfected with SIRT6 and/or PARP1 or PARP1 mutants. Both SIRT6 and PARP1 are required for the stimulation of repair. PARP1 Y889C is a catalytically inactive PARP1. PARP1 DEEKKK contains mutations in all six poly-ADP-ribosylation sites. PARP1 DEEKK contains the same mutations, except at K521. Asterisks indicate values significantly different from control (P < 0.01).
(D) PARP1 K521 is essential for activation of NHEJ by SIRT6. An NHEJ assay was performed in PARP1−/− MEFs containing an integrated NHEJ reporter. Cells were transfected with SIRT6 and/or PARP1 or PARP1 mutants. Both SIRT6 and PARP1 are required for the stimulation of repair. PARP1 Y889C is a catalytically inactive PARP1. PARP1 DEEKKK contains mutations in all six poly-ADP-ribosylation sites. PARP1 DEEKK contains the same mutations, except at K521. Asterisks indicate values significantly different from control (P < 0.01). (E) PARP1 lacking the catalytic domain is mono-ADP-ribosylated by SIRT6 in vitro, whereas K521A is not. (F) In vitro assay of PARP1 poly-ADP-ribosylation activity showing that PARP1 is stimulated only by SIRT6 mono-ADP-ribosylation activity. (G) Stimulation of NHEJ and HR by SIRT6 is abolished by PARP1 inhibitors 3-ABA or PJ34.
Binding of Proteins to DNA Often Involves Hydrogen Bonding Types of domains that bind DNA: Helix-turn-helix Zinc Finger Leucine Zipper Helix-loop-helix
Functional groups on all four base pairs that are displayed in the major and minor grooves of DNA
A = H bond acceptor D = H bond donor
Guanine-Arginine: One of the most common DNA-protein interactions. Because of its specific geometry of H-bond acceptors, guanine can be unambiguously recognized by the side chain of arginine
Helix-Turn-Helix Motif is Common in DNA-Binding Proteins One of the helixes (red) fits into the major groove of DNA Four DNA-binding helix-turn-helix motifs (gray) in the Lac repressor
Important Points: a handshake leads to a bear-hug Specific recognition of DNA targets by the helix-turn-helix motif involves interactions between sides of the recognition helix and bases in the major groove of the DNA But, specific recognition of DNA sequences is to a large extent governed by other interactions within complementary surfaces between the protein and the DNA These interactions frequently involve H-bonds from protein main-chain atoms to the DNA backbone in both the major and the minor groove and are dependent on the sequence-specific deformability of the target DNA
Helix-turn-helix. (a) DNA-binding domain of the Lac repressor The helix-turn-helix motif is shown in red and orange; the DNA recognition helix is red.
Helix-turn-helix. (c) Surface rendering of the DNA-binding domain of the Lac repressor bound to DNA.
The DNA-binding domain separated from the DNA, with the binding interaction surfaces shown. groups on the protein and DNA that interact through H-bonding groups that interact through hydrophobic interactions
Chapter 8: Summary In this chapter, we learned about: • Function of nucleotides and nucleic acids • Names and structures of common nucleotides • Structural basis of DNA function • Reversible denaturation of nucleic acids • Chemical basis of mutagenesis
Hexons and pentons form capsid (TP) Covalently linked to DNA 36 kbp 50 nm Transcribed by RNA pol II Transcribed by RNA pol III Figure A-1 Adenovirus