Download

1 / 42
430 likes | 566 Views
How ubiquitin and SUMOs control trx. Why this?. Transcription. Ubiquitylation of proteins not only targets them for destruction, but is also a regulatory event in the nucleus

E N D
Why this? Transcription • Ubiquitylation of proteins not only targets them for destruction, but is also a regulatory event in the nucleus • In recent years, important connections between ubiquitylation, chromatin structure, signaling pathways and transcriptional control have emerged. • The Ub-proteasome system is ideally suited to controlling the distribution, abundance and activity of components of the transcriptional machinery. The life of a protein Proteolysis
Ubiquitin and the protesome .. A reminder
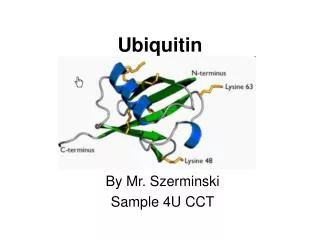
Ubiquitin-family proteins and the proteasome • Ub covalently linked to targets • The ubiquitin (Ub) system defines a family of related modifier proteins that are linked covalently to target proteins. • Ubiquitin is the defining member of this class, • But at least nine other related proteins with this function have been described (see figure). • Degron recognition • Ubiquitylation is a specific process that is signalled by an element — a degradation signal (degron) — in the substrate protein. • The degron is recognized by a Ub-ligase (Ubl), E3,which in turn recruits a Ub-conjugating (Ubc) enzyme, E2, to the substrate. The E3 then catalyses the transfer of Ub groups to a lysine (K) residue that is somewhere in the target protein.
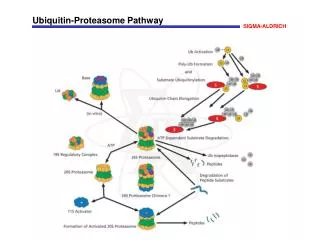
Ubiquitin-family proteins and the proteasome • Multi-Ub = Targeted for proteosomal destruction • The exact nature of ubiquitylation determines the fate of the substrate protein. If a multi-Ub chain — linked by lysine 48 (K48) in Ub itself — forms, the substrate is targeted for destruction by a large, self-compartmentalized, protease known as the 26S proteasome. • The 19S subcomplex of the proteasome recognizes the multi-ubiquitylated substrate, removes the Ub groups, unfolds the substrate and feeds it into the core of the 20S subcomplex where it is destroyed. • If, however, the multi-Ub chain is linked by lysine 63 (K63), or if it has less than four Ub chains, proteolysis does not occur. • A family of Ubi-related small proteins can also be conjugated
Nobel Prize in Chemistry for 2004 • "for the discovery of ubiquitin-mediated protein degradation” • Aaron Ciechanover • Technion – Israel Institute of Technology, Haifa, Israel, • Avram Hershko • Technion – Israel Institute of Technology, Haifa, Israel and • Irwin Rose • University of California, Irvine, USA
Ubiquitylation of histones • Histone H2A and H2B Ubiquitylation - one of the first recognized markers of trx active chromatin • The first ubiquitylated protein to be described was histone H2A • ubiquitylated forms of histones H2A and H2B were associated specifically with actively transcribed genes • Later also H1 and H3 reported to be ubiquitylated
Ubiquitylation and the histone ”code” • Ubiquitylation of chromatin • The ubiquitin (Ub)-conjugating enzyme Rad6 ubiquitylates K123 in the core of histone H2B. This modification promotes the methylation of another histone, H3, at two positions, K4 and K79. These modifications, in turn, are required for telomeric-gene silencing. • TAFII250 (TFIID component) can ubiquitylate the linker histone H1; might relate to the role of this TAF in transcriptional activation. • Ubiquitylation = an integral part of the histone ”code” • Mechanism? • Direct structural role by loosening chromatin structure • Or as ”tag” recognized by proteins such as the proteasome or HDAC6
A coming role for De-ubiquitylation? • Ubps - possible novel regulators? • studies have identified Ub-specific proteases (Ubps) associated with components of both the SIR4 silencing and the SAGA chromatin remodelling complexes.
DNA damage - use of RNAPII to direct repair to active genes • Regulation of trx-coupled repair (TCR) by ubiquitylation of RNA polymerase II. • Transcription-coupled repair (TCR) is the mechanism through which mutations in actively transcribed genes are preferentially repaired. • Elongating RNAPII, with a unique pattern of CTD phosphorylation, encounters a damaged DNA segment. Here the stalled polymerase recruits the Ub-ligase Rsp5, which in turn ubiquitylates the largest subunit of pol II. • Ubiquitylation is followed by the proteasomal destruction of at least one subunit of polymerase, recruitment of the repair machinery and restoration of DNA integrity. • Rsp5 is also a co-activator for the steroid hormone receptors ?
Three strategies • Controlling the localization of the TF • Controlling the activity of the TF • Controlling the abundance of the TF
Regulating TFs by the ubiquitylation - 3 strategies and 4 models • Regulating location. • As with NFkB, the TF can be maintained outside the nucleus by interactions with an inhibitor (IkB) that is destroyed by the Ub–proteasome system. • Another Ub-family member SUMO (S) can directly conjugate to activators and sequester them into nuclear bodies. • Regulating activity. • Ubiquitylation can regulate the association of activators with co-activator proteins either directly, by blocking the association of an activator with its essential cofactor, or indirectly, by facilitating the exchange of cofactors with an activator.
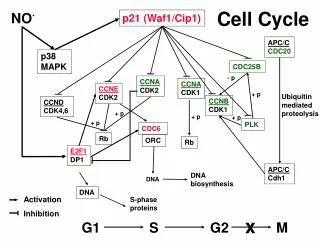
Regulating TF abundance - 1. model • Regulating abundance I - constitutive turnover. • By maintaining an activator in a constitutively unstable form, cells are primed for a transcriptional response when appropriate. In this model, a signal from outside the nucleus leads to a transient stabilization of the activator, which elicits a rapid induction of target genes. • Examples • p53 - later lecture • Wnt-signalling
Destroying TFs when not needed- shutting off proteolysis then gives a rapid response • Beta-catenin and wnt-signalling Signalling Inactivation of GSKb Stabilization of b-catenin Phosphorylation ubiquitylation Degradation Rapid Accumulation To the nucleus
Regulating TF abundance - 2. model • Regulating abundance II - trx-coupled destruction • In this model, activators are destroyed during the act of transcriptional activation as a way of limiting uncontrolled activation by any one DNA-bound transcription factor.
Link: trx activation degradation • TFs often unstable • TAD overlaps closely with degrons • Degron = domain that signals ubiquitination • Myc and many others • Strong activators = rapidly degraded • Weak activators = more stable • Q-rich, N-rich • Mutant TADs with activation lost = stabilized • Link: activation - degradation • TAD ≈ degron • Strong TAD = highly unstable
TADs and degrons overlap • Marking and destroying active TFs are part of into the activation process itself. • A functional relationship between Trx Activation Domain (TADs) and degradation signals (DEGRONS). • The transcriptional activation domains (TADs) and degradation signals (degrons) overlap in 19 unstable transcription factors
Transcriptional activation- risky business? • Evidence indicates that marking and destroying active TFs are part of into the activation process itself. • Kamikaze activators
Srb10 (Cdk8) also targets activator (Gcn4p) sentencing it for destruction CTD Ubiquitinated Degraded
Gcn4 = targeted by Srb10, on the way to destruction • Gcn4 is phosphorylated by Srb10 • Phospho-Gcn4p is recognized by Ub-ligase complex SCFCdc4 • WD40 repeats mediates substrate recognition • Ubiquitinylation of Gcn4p
Basal trx machinery can mark the activators it has encountered, sentencing them to an early death A ”black widow” model • Why should Srb10 destroy the activator? • = Activators are destroyed as a direct consequence of recruiting the basal trx machinery to a promoter
Cyclic processes in trx- role of the Proteasome (Gannon version)
3. conclusion • VP16 TAD signals ubiquitination through Met30 ubi-ligase • Met30 is required for VP16 TAD to activate • This requirement circumvented by Ub-fusion • Activator ubiquitination is essential for trx activation • Ubiquitination = dual signal for activation and activator destruction
A unified model? • In this model, the ubiquitin (Ub)–proteasome system regulates transcription at numerous levels. • Interactions of a TF (activator) with the general trx machinery (green) functions to recruit ubiquitin ligase(s) to the site of transcription and ubiquitylates many factors, including the activator, RNAPII and histones. • These ubiquitylation events in turn recruit the 26S proteasome, which simultaneously destroys the activator and promotes elongation of transcription by pol II.
A unified model? • Limiting uncontrolled trx • Importantly, this proposed mechanism limits uncontrolled transcription in two ways - by destroying the activator at each cycle of promoter ‘firing’ and by ensuring that interactions between pol II and the proteasome are made in an activator- and promoter-dependent manner.
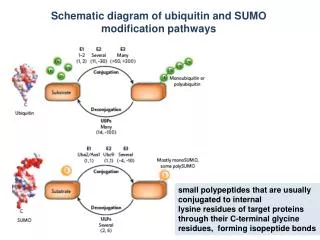
What is SUMO-1 ? • SUMO-1 (small ubiquitin-related modifier) • peptide of 101 residues / mature polypeptide 98 residues • function ≠ ubiquitin • NOT tagged for degradation • Rather stabilized or ”targeted” to sub-nuclear structures
The SUMO-1 protein • Small Ubiquitin MOdifier • Link: isopeptide bond • between the C-terminal glycine of SUMO and the e-amino group of a lysine residue in the target protein. • Structure • characteristic ubiquitin-fold + unique unstructured N-terminal extension of up to 22 residues - possible protein interaction site? GG XKE
Many nuclear targets • Largely nuclear target proteins • NLS + KxE • A short peptide that contains the KxE motif and a NLS suffices to produce a SUMO conjugate in vivo. • Mutated NLS abolish Sumoylation • SP100, HDAC4, MDM2
Sumoylation - functional roles • Antagonizing other modifications • SUMO modification of IkBa stabilizes this NF-kB inhibitor by blocking ubiquitylation at the same acceptor site. • Confer new interactions • Conformational change • New interaction surface – enhance or inhibit interactions • Altering the subcellular localization of the protein • Sumoylation causes the relocalization of the nuclear import factor RanGAP1 from the cytoplasm to the nuclear pore complex (NPC). • Many Trx factors associated with PML nuclear bodies
Sumoylation – co-repressor recruitment • Sumoylated TF promotes binding of co-repressor.
Sumoylation and nuclear import • NPC passage • A substrate that contains a NLS might be sumoylated at the nuclear pore by the E3 ligase activity of RanBP2, after which it might be de-modified by a Ulp1-type SUMO protease that resides at the nucleoplasmic face of the nuclear pore complex (NPC), or by a Ulp2-type, nucleoplasmic protease. • Nuclear dynamic modification • Once inside the nucleus, substrates might undergo SUMO modification that is mediated by PIAS or Pc2 E3 ligases.
Zhong et al. (2000) Nature Cell Biol. 2:E85-E90 PML and PML nuclear bodies - SUMO required
Regulating the trx InitiationMachinery by Lysine Modification