Download

1 / 9
100 likes | 463 Views
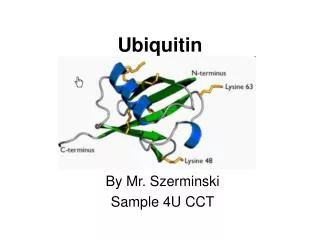
Ubiquitin and Ub-like proteins. 18-1. Ubiquitin and ubiquitin-like proteins - background - the ubiquitin fold - ThiS, molybdopterin synthase Degradation - degradation of newly-synthesized proteins. Ubiquitin. ubiquitin is always made as a fusion protein:

E N D
Ubiquitin and Ub-like proteins 18-1 Ubiquitin and ubiquitin-like proteins - background - the ubiquitin fold - ThiS, molybdopterin synthase Degradation - degradation of newly-synthesized proteins
Ubiquitin • ubiquitin is always made as a fusion protein: - ubiquitin cleaved after GlyGly by enzyme • Ubiquitin-like proteins were first identified and characterized ~5 years ago N C GG Ubiquitin fusion protein (often a ribosomal subunit) 18-2 • Schlesinger and Goldstein (1975) “Molecular conservation of the 74 amino acid sequence of ubiquitin between cattle and man”Nature255, 42304. • note: it is actually 76 amino acids in length • Wolf et al. (1993) “Ubiquitin found in the archaebacterium Thermoplasma acidophilum”FEBS Lett.326, 42. • Sequencing of the first bacteria and archaea (1995/1996): no sign of ubiquitin! • ubiquitin is the most highly conserved protein in eukaryotes and is not found in prokaryotes • how can such a protein arise in eukaryotes only? Is there not an ancestral ubiquitin-like protein in prokaryotes? • ubiquitinated proteins are recognized and degraded by the 26S proteasome in eukaryotes after a complex ubiquitination pathway - archaea possess a proteasome as well (20S) but no regulatory particle which recognizes ubiquitin
Ubiquitin-fold structures ubiquitin Nedd8 UBX ThiS 18-3 • ubiquitin, Nedd8, UBX and ThiS all have ubiquitin folds (i.e., similar structures) • ThiS is the most divergent, with only 14% sequence identity with ubiquitin; in comparison, Nedd8 is >50% identical • all of the sequences contain C-terminal Gly-Gly residues that is used for conjugation; this Gly-Gly terminus is perfectly conserved
ThiS 18-4 • ThiS is a sulfur ‘carrier’ protein that plays a central role in thiamin biosynthesis in E. coli • during thiamin biosynthesis, sulfur from Cysteine is transferred to thiazole, which is then incorporated into thiamin O O ThiF ThiF ThiS ThiS ThiS ATP Cys AMP SH ThiF, ThiH, Thil, ThiG S OP OP S + N N N thiazole thiamin reaction scheme for the biosynthesis of thiamin N NH2
Molybdopterin synthase 18-5 • Molybdopterin (MPT) synthase is an evolutionarily-conserved enzyme that is present in bacteria, archaea and eukaryotes; it is a dimeric protein that consists of MoaD and MoaE, and together with MoeB catalyzes the formation of Molypdopterin • Molypdopterin plays a role in sulfite detoxification and the metabolism of xenobiotics • defects in molydopterin biosynthesis results in human disease OH O MoaE O P OH O O precursor z MPT synthase (MoaD, MoaE) and MoeB MoaD (Ub-like fold) ubiquitin for comparison SH SH OPO3- O molybdopterin
Ubiquitin/Ub-like activation pathways *ThiF* AMP S-ThiF Gly C Gly C O (2) Gly C ThiS O O O ATP PPi S-MoeB Gly C O AMP (3) Gly C Gly C *MoeB* MoaD O O O ATP PPi SH Gly C Thil, cys (2) thiamin O SH Gly C X-S Y (3) molybdopterin O 18-6 • all proteins in ubiquitin superfamily are used in a very similar activation pathway, with the activating enzyme being homologous (1) Uba1 (E1) E1 O AMP S-E1 Gly C Gly C Gly C ubiquitin O O O ATP PPi multi- Ub’d protein S-E2 NH-protein Gly C Gly C E2 E3 (1) O O
transports iron JAB domain: related to de-ubiquitinating isopeptidase
Degradation of newly-synthesized proteins 18-7 First: one important use of proteasome degradation products • proteins are degraded down to 8-10-mer peptides by the proteasome • the peptides are then imported into the ER by the peptide transporter associated with antigen processing (TAP) • MHC class I receptors containing bound peptides are then presented on the cell surface
DRiPs *Schubert et al. (2000) Rapid degradation of a large fraction of newly synthesized proteins by proteasomes. Nature 404, 770-774. 18-8 DRiPs, Defective Ribosomal products • DRiPs represent polypeptides that never attain native structure owing to errors in translation or post-translational processes necessary for the proper biogenesis of the proteins • Schubert et al.* found that upwards of 30% of all newly-synthesized proteins from various cell types are degraded by the proteasome • at least some of the DRiPs are ubiquitinated • a ubiquitinated DRiP is formed from HIV Gag polyprotein, a long-lived viral protein that serves as a source of antigenic peptides • presentation of MHC class I molecules require continuing protein translation, implying that peptides that are presented are derived from newly-made proteins - does it make sense that the cell should degrade 30% of its proteins before they have a chance to function? - might the degradation of a large fraction of DRiPs represent a defence mechanism against viral intrusions?