Download
1 / 79
800 likes | 1.26k Views
Bioquímica Estructural Lípidos y mem branas biológicas. 8.1. Introducción . 8.2. Funciones de los lípidos. Función energética. Función de reserva de agua. Función de producción de calor. Función estructural. Función como hormonas y segundos mensajeros. Función como vitaminas.

E N D
8.1. Introducción. 8.2. Funciones de los lípidos. • Función energética. • Función de reserva de agua. • Función de producción de calor. • Función estructural. • Función como hormonas y segundos mensajeros. • Función como vitaminas. 8.3. Clasificación de los lípidos. • Lípidos saponificables. • Lípidos no saponificables. • Otros lípidos. 8.4. Lípidos saponificables. • Ácidos grasos saturados, insaturados y derivados. • Eicosanoides: prostaglandinas, tromboxanos y leucotrienos. • Lípidos neutros: acilgliceroles y ceras. • Lípidos anfipáticos: glicerolípidos y esfingolípidos. 8.5. Lípidos no saponificables. • Terpenos: • Retinoides. • Carotenoides. • Tocoferoles. • Naftoquinonas. • Dolicoles. • Esteroides: • Ácidos y sales biliares. • Esteroles. • Hormonas esteroideas: estrógenos, andrógenos, gestágenos y corticoides. 8.6. Otros lípidos. • Lípidos pirrólicos de cadena abierta y cerrada. • Hidrocarburos. 8.7. Trabajo con lípidos en el laboratorio. • Extracción de lípidos en disolventes orgánicos. • Cromatografía de absorción: TLC. • Cromatografía gás líquido. • Espectrometría de masas.
Bibliografía Los contenidos de este tema pueden ser ampliados en: Capítulo 12 de Berg, J. M., Tymoczko, J. L. y Stryer, L. "Bioquímica", 6a ed. 2008 Capítulos 10 de Nelson, D. y Cox, M. "Lehninger, Principios de Bioquímica", 4ªed. 2006. Los objetivos específicos pueden encontrarse en la guía docente de la asignatura. Los artículos y libros específicos relativos a este tema pueden encontrarse en la página web: shaker.umh.es
8.1. Introducción. Denominamos lípidos a un conjunto muy heterogéneo de biomoléculas cuya característica distintiva, aunque no exclusiva ni general, es su insolubilidad en agua, y su solubilidad en disolventes orgánicos (benceno, cloroformo, éter, hexano, etc.). Los lípidos pueden encontrarseunidos covalentemente con otras biomoléculas como en el caso de los glicolípidos (presentes en las membranas biológicas), las proteínas aciladas (unidas a algún ácido graso) o las proteínas preniladas (unidas a lípidos de tipo isoprenoide). También forman asociaciones no covalentes con otras biomoléculas, como en el caso de las lipoproteínas y de las estructuras de membrana. Una característica básica de los lípidos, y de la que derivan sus principales propiedades biológicas es la hidrofobicidad. La baja solubilidad de los lipídos se debe a que su estructura química es fundamentalmente hidrocarbonada, es decir, con gran cantidad de enlaces C-H y C-C.
8.1. Introducción. El agua, al ser una molécula muy polar, con gran facilidad para formar puentes de hidrógeno, no es capaz de interaccionar con los lípidos. Adopta en torno a ellos una estructura muy ordenada que maximiza las interacciones entre las propias moléculas de agua, forzando a la molécula hidrofóbica al interior de una estructura en forma de jaula, que también reduce la movilidad del lípido. Todo ello supone una configuración de baja entropía, que resulta energéticamente desfavorable. Esta disminución de entropía es mínima si las moléculas lipídicas se agregan entre sí, e interaccionan mediante fuerzas de corto alcance, como las fuerzas de van der Waals. Este fenómeno recibe el nombre deefecto hidrofóbico.
8.2. Funciones de los lípidos. Dado que los lípidos presentan estructuras muy diversas, las funciones biológicas que ejercen son también muy variadas. En muchos organismos las grasas y aceites son las formas principales de almacenamiento energético (Tema 23), mientras que fosfolípidos y esteroles constituyen los principales elementos estructurales de las membranas biológicas (Tema 9). Otros lípidos, presentes en pequeñas cantidades, juegan papeles esenciales. Entre estos últimos podemos destacar papeles como: • Cofactores enzimáticos. • Transportadores electrónicos. • Pigmentos que absorben la luz. • Sitios hidrofóbicos de unión a proteínas. • "Chaperonas" que ayudan al plegamiento de proteínas de membrana. • Agentes emulsionantes en el tracto digestivo. • Hormonas. • Mensajeros intracelulares.
8.2. Funciones de los lípidos: función energética. Los lípidos (generalmente en forma de triacilgiceroles) constituyen la reserva energética de uso diferido del organismo, en comparación con los azúcares, que són de movilización inmediata. Su contenido calórico es muy alto (10 Kcal/gramo), y representan una forma compacta y anhidra de almacenamiento de energía (Tema 23). A diferencia de los hidratos de carbono, que pueden metabolizarse en presencia o en ausencia de oxígeno, los lípidos sólo pueden metabolizarse aeróbicamente (Tema 23).
8.2. Funciones de los lípidos: función de reserva de agua metabólica. Aunque parezca paradójico, los lípidos representan una importante reserva de agua. Al poseer un grado de reducción mucho mayor que el de los hidratos de carbono, su metabolismo aerobio produce una gran cantidad de agua (agua metabólica). Así, la oxidación completa de un mol de ácido palmítico puede producir hasta 146 moles de agua. En animales como camellos y dromedarios, las reservas grasas de su joroba se utilizan principalmente para producir agua.
8.2. Funciones de los lípidos: función de producción de calor. En algunos animales hay un tejido adiposo especializado que se llama grasa parda o grasa marrón. En este tejido, la combustión de los lípidos está desacoplada de la fosforilación oxidativa, por lo que no se produce ATP, y la mayor parte de la energía se disipa en forma de calor. En los animales que hibernan, la grasa parda permite generar la energía necesaria para el animal. Por ejemplo, un oso puede llegar a perder hasta el 20% de su masa corporal.
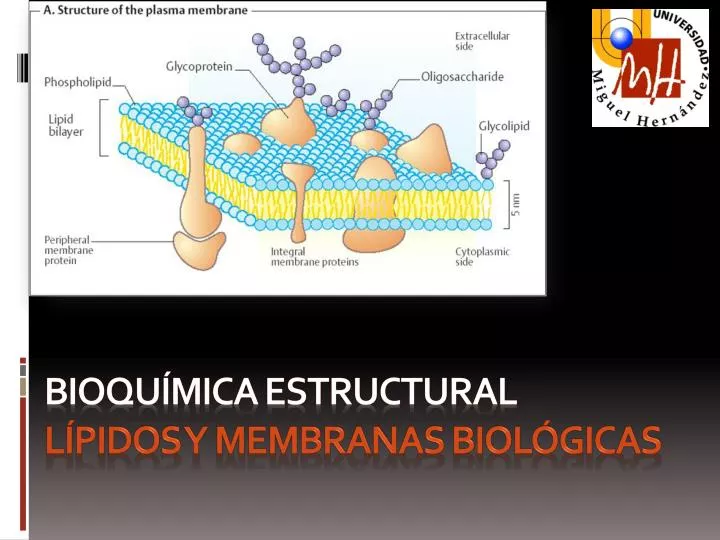
8.2. Funciones de los lípidos: lípidos estructurales de las membranas biológicas. Todas las células y sus orgánulos subcelulares están delimitados por una membrana biológica. Como veremos en el Tema 9, las membranas están formadas por una doble capa lipídica y una compleja batería proteíca, distinta en cada tipo celular y en cada orgánulo subcelular. Las membranas constituyen unas "barreras semipermeables" al paso de moléculas polares e iones. Los lípidos de las membranas son anfipáticos, o sea, poseen un extremo de la molécula hidrofóbico y otro hidrofílico. Las interacciones hidrofóbicas entre ellos y las hidrofílicas con el agua permiten su empaquetamiento formando estas bicapas.
8.2. Funciones de los lípidos: lípidos estructurales de las membranas biológicas. Se describirán cinco tipos generales de lípidos de membrana: Glicerofosfolípidos, cuya región hidrofóbica consta de dos ácidos grasos unidos al glicerol. Galactolípidos y sulfolípidos, que también tienen dos ácidos grasos esterificando el glicerol pero carecen del fosfato típico de los fosfolípidos. Glicerol dialquil glicerol tetraéteres, presentes en las arqueobacterias y que presentan dos largas cadenas alquílicas unidas por enlace éter al glicerol en ambos extremos. Esfingolípidos, en los que un ácido graso se une a la esfingosina por enlace amida. Esteroles, que presentan un sistema rígido de 4 anillos hidrocarbonados fusionados.
8.2. Funciones de los lípidos: función como hormonas. Los organismos pluricelulares han desarrollado distintos sistemas de comunicación entre sus órganos y tejidos. En ellos, el sistema endocrino genera señales químicas para la adaptación del organismo a circunstancias diversas. Estas señales reciben el nombre de hormonasy muchas de ellas son lípidos (esteroides, prostaglandinas, leucotrienos, calciferoles, etc).
8.2. Funciones de los lípidos: función como segundos mensajeros intracelulares. En otros casos, los lípidos pueden funcionar como segundos mensajeros intracelulares. Esto ocurre cuando se activan las fosfolipasas o las esfingomielinasas que hidrolizan glicerolípidos o esfingolípidos generando diversos compuestos que actúan como segundos mensajeros. Como ejemplos tenemos diacilgliceroles, ceramidas, inositolfosfatos, etc. que intervienen en muchos procesos celulares.
8.2. Funciones de los lípidos: función como cofactores enzimáticos. Hay una serie de sustancias que son vitales para el correcto funcionamiento del organismo, y que no pueden ser sintetizadas por éste. Por ello deben ser suministradas en su dieta. Estas sustancias reciben el nombre de vitaminas. La función de muchas vitaminas consiste en actuar como cofactores de enzimas (Temas 10, 11 y 12).
8.3. Clasificación de los lípidos: saponificables y no saponificables. La heterogeneidad estructural de los lípidos dificulta su clasificación sistemática. El componente lipídico de una muestra biológica puede ser extraído con disolventes orgánicos y sometido a un criterio empírico para clasificar sus componentes, este criterio es la reacción de saponificación. La saponificación consiste en una hidrólisis alcalina de la preparación lipídica (con KOH o NaOH). De esta reacción resultan lípidos derivados de ácidos grasos dan lugar a sales alcalinas (jabones) y alcoholes. No todos los lípidos presentes en una muestra biológica dan lugar a este tipo de reacción.
8.4.1. Lípidos saponificables: ácidos grasos saturados, insaturados y derivados. Los ácidos grasosson ácidos monocarboxílicos de cadena larga. Por lo general, contienen un número par de átomos de carbono, normalmente entre 12 y 24. Ello se debe a que su síntesis biológica tiene lugar mediante la aposición sucesiva de unidades de dos átomos de carbono. Sin embargo, también existen ácidos grasos con un número impar de átomos de carbono, que probablemente derivan de la metilación de un ácido graso de cadena par.
8.4.1. Lípidos saponificables: ácidos grasos saturados, insaturados y derivados. Laspropiedades químicasde los ácidos grasos derivan por una parte, de la presencia de un grupo carboxilo, y por otra parte de la existencia de una cadena hidrocarbonada. La coexistencia de ambos componentes en la misma molécula, convierte a los ácidos grasos en moléculasdébilmente anfipáticas (el grupo COOH es hidrofílico y la cadena hidrocarbonada es hidrofóbica). La solubilidad en agua decrece a medida que aumenta la longitud de la cadena.
8.4.1. Lípidos saponificables: ácidos grasos saturados, insaturados y derivados. El grupo carboxílico de la molécula convierte al ácido graso en un ácido débil (con un pKa en torno a 4,8). También presenta las reacciones químicas propias del grupo COOH: esterificación con grupos OH, formación de enlaces amida con grupos NH2, formación de sales (jabones), etc. El grupo COOH es capaz de formar puentes de hidrógeno, de forma que los puntos de fusión de los ácidos grasos son mayores que los de los hidrocarburos correspondientes.
8.4.1. Lípidos saponificables: ácidos grasos saturados, insaturados y derivados. Según la naturaleza de la cadena hidrocarbonada, distinguimos tres grandes grupos de ácidos grasos: Ácidos grasos saturados. Ácidos grasos insaturados. Derivados de ácidos grasos: • jabones, • hidroxiácidos grasos, • ácidos grasos ramificados, • ácidos grasos cíclicos, • ácidos grasos con triples enlaces. • Eicosanoides: • Prostaglandinas. • Tromboxanos. • Leucotrienos.
8.4.1. Lípidos saponificables: ácidos grasos saturados. Desde el punto de vista químico, son muy poco reactivos. Por lo general, contienen un número par de átomos de carbono. En la nomenclatura de los ácidos grasos se utilizan con más frecuencia los nombres vulgares que los sistemáticos. La nomenclatura abreviadaes muy útil para nombrar los ácidos grasos. Ejemplo, C18:0
8.4.1. Lípidos saponificables: ácidos grasos saturados. Los ácidos grasos saturados más abundantes son el palmítico (C16:0) y el esteárico (C18:0). Los ácidos grasos saturados de menos de 10 átomos de C son líquidos a temperatura ambiente y parcialmente solubles en agua. A partir de 12 C, son sólidos y prácticamente insolubles en agua. El punto de fusión aumenta con la longitud de la cadena. Los ácidos grasos de cadena imparprobablemente derivan de la metilación de un ácido graso de cadena par. Ejemplos son el valeriánico (C5:0) y pelargónico (C9:0).
8.4.1. Lípidos saponificables: ácidos grasos insaturados. Con frecuencia, aparecen insaturaciones en los ácidos grasos, mayoritariamente en forma de dobles enlaces, aunque se han encontrado algunos con triples enlaces. Cuando hay varios dobles enlaces en la misma cadena, estos no aparecen conjugados(alternados), sino cada tres átomos de carbono. La posición de los dobles enlaces se indica como un superíndice en el segundo número después de la letra ∆. Ej. Ácido linolénico C18:3 ∆ 9,12,15.
8.4.1. Lípidos saponificables: ácidos grasos insaturados. Por lo general, las insaturaciones de los ácidos grasos son del tipo cis. Esto hace que la disposición de la molécula sea angulada, con el vértice en la insaturación. Los ácidos insaturados tienen puntos de fusión más bajos que los de sus homólogos saturados. Los dobles enlaces en trans distorsionan poco la simetría cristalina, que es muy parecida a la de los ácidos grasos saturados. Ejem. ácido araquidónico se representa como C20:45c,8c,11c,14c.
8.4.1. Lípidos saponificables: ácidos grasos insaturados. Algunos ácidos grasos poliinsaturados (linoleico, linolénico y araquidónico) no pueden ser sintetizados por los animales superiores (incluído el hombre), y como su función biológica es fundamental, deben ser suministrados en la dieta. Por este motivo reciben el nombre de ácidos grasos esenciales. Los ácidos grasos insaturados manifiestan las propiedades inherentes al doble enlace: - Reaccionan con ácido sulfúricopara dar sulfonatos, que se emplean como detergentes domésticos. - Pueden adicionar hidrógeno. La hidrogenación catalítica de los ácidos grasos insaturados constituye la base de la transformación industrial de aceites en grasas sólidas (la margarina es el resultado de la hidrogenación de aceites vegetales). - Pueden autooxidarse con el oxígeno del aire, contribuyendo al fenómeno de enranciamiento de las grasas de los alimentos almacenados.
8.4.1. Lípidos saponificables: tabla resumen sobre ácidos grasos.
8.4.1. Lípidos saponificables: derivados de ácidos grasos. JABONES: son las sales de los ácidos grasos. Debido a la polaridad del anión carboxilato tienen un fuerte carácter anfipático, y son muy miscibles con el agua, especialmente los jabones de metales alcalinos. En general, los jabones adoptan en medio acuoso estructuras micelares en equilibrio con formas libres. Las grandes micelas esféricas pueden incluir en su interior grasas neutras, por lo que los jabones tienen poder detergente.
8.4.1. Lípidos saponificables: derivados de ácidos grasos. HIDROXIÁCIDOS GRASOS: contienen grupos hidroxilo en la cadena hidrocarbonada. Ejemplos son el ácido cerebrónico(2-hidroxi C24:0), el hidroxinervónico (2-hidroxi C24:1∆ 15), ambos presentes en esfingolípidos de cerebro. Aceit e
8.4.1. Lípidos saponificables: derivados de ácidos grasos. ÁCIDOS GRASOS RAMIFICADOS: contienen uno o varios grupos metilo como sustituyentes en la cadena hidrocarbonada. Ejem. el ácido tuberculoesteárico(10-metil C18:0), presente en el bacilo de la tuberculosis (Mycobacterium tuberculosis). En el hombre, el ácido fitánicoaparece como consecuencia de deficiencias en el metabolismo del fitol (un componente de la molécula de clorofila), que no puede ser degradado en el hígado.
8.4.1. Lípidos saponificables: derivados de ácidos grasos. ÁCIDOS GRASOS CÍCLICOS: contienen un anillo de ciclopropano (ácido lactobacílico) o de ciclopentano (ác. chaulmógrico) que se encuentra en semillas de plantas. ÁCIDOS GRASOS CON TRIPLES ENLACES: algunos actúan como antibióticos (Ej. ácido nemotínico).
8.4.2. Lípidos saponificables: eicosanoides. Todos los eicosanoides proceden del ácido araquidónico, un ácido graso insaturado de 20 carbonos, C20:4 5c,8c,11c,14c. Los eicosanoides acúan como hormonas paracrinas, es decir, sustancias que actúan sólo en células próximas al punto de síntesis, en lugar de ser transportadas por la sangre para actuar en células de otros tejidos u órganos. Ejercen una gran variedad de efectos sobre diversos tejidos de vertebrados. Hay tres clases de eicosanoides: • Prostaglandinas. • Tromboxanos. • Leucotrienos.
8.4.2. Lípidos saponificables: eicosanoides del tipo prostaglandinas. Las protaglandinas contienen un anillo de 5 átomos de carbono. Fueron originalmente aisladas de la glándula prostática. Existen varias familias de PG, que se denominan con una letra adicional (PGA, PGB, PGC, PGD, PGE, PGF, etc), en función de los sustituyentes del anillo ciclopentano de su estructura. A menudo, la letra mayúscula va seguida de un subíndice que indica el número de dobles enlaces presentes en la molécula, sin incluir el anillo. Se conocen unas 20 PG, y sus funciones son muy variadas sobre distintos tejidos. Las PGE y PGF provocan la contracción de la musculatura lisa, en especial en el aparato reproductivo. La PGI2 es un vasodilatador que actúa principalmente sobre las arterias coronarias y que impide la agregación plaquetaria. Las PGG y PGH son mediadores de la reacción inflamatoria. Compuestos como el ácido acetilsalicílico (aspirina) inhiben la síntesis de estas PG, y de ahí sus efectos antiinflamatorios.
8.4.2. Lípidos saponificables: eicosanoides del tipo tromboxanos. Los tromboxanos contienen un anillo de 6 átomos que contiene una función éter. Son producidos por las plaquetas y actúan en la formación de coágulos sanguíneos y en la reducción del flujo sanguíneo hacia el lugar del coágulo. Los antiinflamatorios no esteroideos (aspirina, ibuprofeno y meclofenamato) inhiben a la enzima protaglandina H2 sintasa, que cataliza uno de los primeros pasos del ácido araquidónico hacia prostaglandinas y tromboxanos.
8.4.2. Lípidos saponificables: eicosanoides del tipo leucotrienos. Los leucotrienos fueron inicialmente descritos a partir de leucocitos. Contienen 3 dobles enlaces conjugados. Como ejemplo tenemos el LA4 que induce la contracción del músculo que recubre la vias del pulmón. Su sobreproducción conlleva ataques asmáticos, de forma que su síntesis constituye una de las dianas de los medicamentos antiasmáticos.
8.4.3. Lípidos neutros: ceras y acilgliceroles. Son ésteres de ácidos grasos con alcoholes. No tienen ningún otro tipo de componentes, por lo que son moléculas muy poco reactivas. En la Naturaleza encontramos dos tipos: • Ceras • Acilgliceroles Las ceras biológicas son ésteres de ácidos grasos de cadena larga saturados o insaturados (de 14 a 36 átomos de carbono) con alcoholes de cadena larga (16 a 30 átomos). Sus puntos de fusión, en 60 y 100 ºC, son más altos que los de los triacilgliceroles. En el plancton marino las ceras son la principal forma de almacenamiento de combustible metabólico.
8.4.3. Lípidos neutros: ceras y acilgliceroles. Ciertas glándulas de la piel de vertebrados secretan ceras para proteger el pelo y la piel manteniéndolos flexibles, lubricados e impermeables. Las aves secretan ceras para impermeabilizar sus plumas. Las hojas de acebo y de muchas plantas tropicales secretan ceras para protegerse de parásitos e impedir una excesiva evaporación del agua. Diversas ceras biológicas (como la lanolina, la cera de abeja o el aceite de espermaceti del cachalote) son ampliamente usadas por la industria farmaceútica y cosmética.
8.4.3. Lípidos neutros: ceras y acilgliceroles. Los acilgliceroles o glicéridos son ésteres de ácidos grasos con glicerol(propanotriol). Constituyen el contingente mayoritario de los lípidos de reserva energética, y son muy abundantes en el tejido adiposo animal y en las semillas y frutos de las plantas oleaginosas. El glicerolpresenta tres grupos alcohólicos, y por tanto puede aparecer esterificado en una, dos o tres posiciones, dando lugar respectivamente, amonoacilgliceroles, diacilgliceroles ytriacilgliceroles.
8.4.3. Lípidos neutros: ceras y acilgliceroles. Los adipocitos de vertebrados almacen triacilgliceroles en forma de gotas de grasa que ocupan casi toda la célula. De igual forma muchas semillas de plantas también almacenan triacilgliceroles que proporcionan energía y precursores biosintéticos durante la germinación. Tanto adipocitos como semillas germinadas poseen lipasas que hidrolizan los triacilgliceroles y dan lugar a ácidos grasos.
8.4.4. Lípidos anfipáticos. Los lípidos anfipáticos se pueden clasificar en función de la naturaleza del alcohol al que se encuentran unidos los ácidos grasos, y distingue dos grandes grupos: Los glicerolípidos, en los que los ácidos grasos están esterificados a los carbonos sn-1 y sn-2 del glicerol, Los esfingolípidos, en los que el ácido graso se une a la esfingosina, un alcohol nitrogenado de 18 átomos de carbono por un enlace amida.
8.4.4. Lípidos anfipáticos: glicerolípidos. Los glicerolípidos son lípidos anfipáticos en los que está presente glicerol, al que se unen: Dos ácidos grasos por sendos enlaces éster , además de un aminoalcohol a través de un enlace fosfodiester(glicerofosfolípidos). Algún alqueno o alcano unido por enlace éter , además de un aminoalcohol a través de un enlace fosfodiester (ej. plasmalógenos). Dos ácidos grasos por sendos enlaces éster , además de galactosa o sulfogalactosa a través de un enlace glucosídico (ej. Galactolípidos y sulfolípidos). Dos ácidos grasos largos y ramificadosunidos por sus dos extremos a dos moléculas de glicerol a través de enlaces éter (ej. Glicerol dialquil glicerol tetraéteres de las arqueobacterias). Estos cuatro tipos de glicerolípidos forman parte de las membranas biológicas y todos ellos son moléculas anfipáticas. A continuación describiremos la estructura de cada uno de estos 4 tipos de lípidos anfipáticos.
8.4.4. Lípidos anfipáticos: tipo 1, glicerofosfolípidos. Los glicerofosfolípidos son lípidos de membrana en los que dos ácidos grasos están unidos por enlace éster al primer y segundo carbonos del glicerol, además de un grupo de cabeza polar o cargada unido por un enlace fosfodiester al tercer carbono. La naturaleza de este grupo de cabeza polar define los distintos tipos de glicerofosfolípidos: PA (ácidos fosfatídicos), PC (fosfatidil colinas), PG (fosfatidilgliceroles), PE (fosfatidiletanolaminas), PS (fosfatidilserinas), PI (fosfatidilinositoles) o cardiolipinas. Los ácidos grasos de los glicero-fosfolípidos pueden ser muy variados, ello da lugar a distintas especies moleculares, y por eso hablamos p.ej. de fosfatidilcolinas, en plural. Los ácidos grasos más frecuentes en el C-1 tienen 16 o 18 carbonos y son saturados, mientras en el C-2 es frecuente encontrar un ácido graso insaturado de entre 18 y 20 carbonos.
8.4.4. Lípidos anfipáticos: tipo 1, glicerofosfolípidos. Las enzimas capaces de hidrolizar este tipo de lípidos son las fosfolipasas. Se distinguen varios tipos: La fosfolipasa A1: hidroliza el ácido graso en posición C-1. Genera un lisoglicerolípido y un ácido graso. La fosfolipasa A2: hidroliza el ácido graso en posición C-2. Genera un lisoglicerolípido y un ácido graso. La fosfolipasa C : genera diacilglicerol por un lado y la cabeza polar (con el fosfato) por otro. La fosfolipasa D: Genera ácido fosfatídico por un lado y la cabeza polar (sin el fosfato) por otro.
8.4.4. Lípidos anfipáticos: tipo 2, ejem. plasmalógenos. En algunos tejidos animales y organismos unicelulares podemos encontrar lípidos en los que una de las dos cadenas de ác. graso está unida al glicerol por enlace éter. La cadena unida por enlace éter puede ser saturada, o tener doble enlace entre C-1 y C-2, como ocurre en los plasmalógenos. El tejido cardiaco de vertebrados es rico en plasmalógenos, al igual que las membranas de ciertas bacterias y microorganismos. Se desconoce la importancia de estos lípidos en las membranas, pero está claro que son resistentes a las fosfolipasas.
8.4.4. Lípidos anfipáticos: tipo 3, glicoglicerolípidos. En la membrana tilacoidal interna de cloroplastos de plantas encontramos lípidos en los que uno o dos residuos de galactosa están unidos por un enlace glucosídico con el C-3 del 1,2-diacilglicerol. Estos galactolípidos son probablemente los lípidos de membrana más abundantes de la biosfera. En membranas vegetales también encontramos sulfolípidos, en los que un residuo de glucosa sulfonadose une por un enlace glucosídicoal C-3 del 1,2-diacilglicerol.
8.4.4. Lípidos anfipáticos: tipo 4, glicerol dialquil glicerol tetraéteres de arqueobacterias. La mayoría de las arqueobacterias viven en condiciones extremas (alta temperatura, bajo pH o alta fuerza iónica). En su membrana se han descrito lípidos que contienen largas cadenas hidrocarbonadas ramificadas unidas en cada extremo al glicerol por enlaces éter, que son más estables a las condiciones extremas. Unidos a los gliceroles de ambos extremos se encuentran moléculas de glicerol-3-P y secuencias de oligosacáridos. Genéricamente se conoce a estos lípidos como glicerol dialquil glicerol tetraéteres.
8.4.4. Lípidos anfipáticos: esfingolípidos. Los esfingolípidos están formados por una molécula del amino-alcohol esfingosina o alguno de sus derivados, una molécula de ácido graso unido al C-2 por enlace amida y una cabeza polar unida por enlace glucosídico en unos casos y por enlace fosfodiester en otros, al C-1. La unión por enlace amida del ácido graso a la esfingosina da lugar a una molécula de ceramida. Las ceramidas suelen contener ácidos grasos de cadena muy larga, como el lignocérico, el nervónico o cerebrónico.
8.4.4. Lípidos anfipáticos: esfingolípidos. Hay tres subclases de esfingolípidos y todas derivan de las ceramidas. Se diferencian en sus grupos de cabeza: Esfingomielinas, Glucoesfingolípidos neutros, Gangliósidos. Las esfingomielinas contienen fosfocolina o fosfoetanolamina como grupo de cabeza polar unido al C-1 de la esfingosina por enlace éster. Son moléculas estructuralmente análogas a los glicerofosfolípidos.
8.4.4. Lípidos anfipáticos: esfingolípidos. Los glucoesfingolípidos neutros tienen uno o más azúcares en su cabeza polar unidos al C-1 de la esfingosina a través de un enlace glucosídico. En esta subclase de esfingolípidos diferenciamos a su vez dos subtipos: cerebrósidos, que tienen un único azúcar (glucosa o galactosa). Globósidos, que contienen 2 o más azúcares, normalmente glucosa, galactosa o N-acetil-galactosamina.
8.4.4. Lípidos anfipáticos: esfingolípidos. La tercera subclase de esfingolípidos es la de los gangliósidos y que estructuralmente resultan ser los más complejos. Sus grupos polares de cabeza están formados por oligosacáridos en los que hay uno o varios residuos de ácido N-acetilneuramínico (neu5Ac) . Este ácido aporta carga al gangliósido a pH 7. Los gangliósidos con un residuo de ácido siálico forman la serie GM (M de mono-), los que tienen dos forman la serie GD y así sucesivamente.
8.5. Lípidos no saponificables. isopreno Los lípidos no saponificables más importantes son derivados por aposición de varias unidades isoprénicas, y se sintetizan a partir de una unidad básica de 5 átomos de carbono: el isopreno. En este grupo de lípidos se incluyen: Terpenos: retinoides (vitamina A), carotenoides (provit. A), tocoferoles (vit. E), naftoquinonas (vit. K) y dolicoles. Esteroides: esteroles, sales y ácidos biliares, hormonas esteroideas. Existen otros lípidos no saponificables que no están relacionados estructuralmente con el isopreno: Lípidos pirólicos. Hidrocarburos.