Download
1 / 37
380 likes | 565 Views
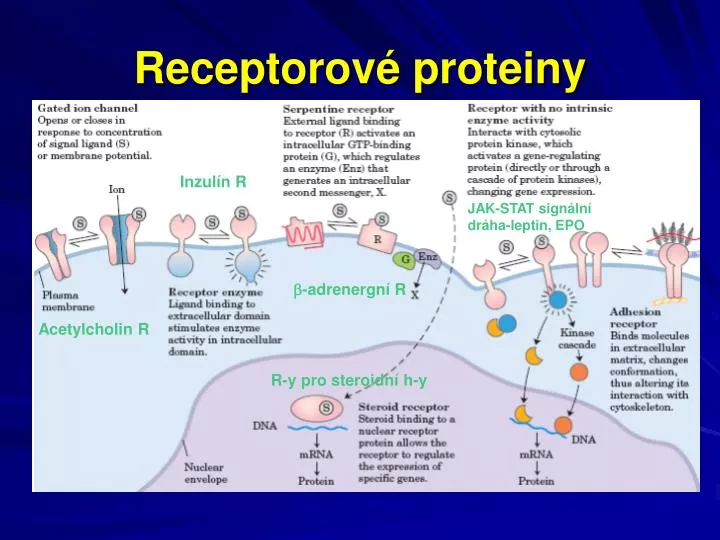
Receptorové proteiny. Inzulín R. JAK-STAT signální dráha-leptin, EPO. -adrenergní R. Acetylcholin R. R-y pro steroidní h-y. JAK-STAT signalizační dráha (receptor EPO, leptinový R). Leptin. Proteinový hormon produkovaný adipocyty

E N D
Receptorové proteiny Inzulín R JAK-STAT signální dráha-leptin, EPO -adrenergní R Acetylcholin R R-y pro steroidní h-y
Leptin • Proteinový hormon produkovaný adipocyty • Kódovaný ob genem (7q31, 3 exony, 2 introny, 15 kbp) • 167 aa (z toho 21 signální sekvence) 146 aa • Známo 5 izoforem leptinového R-u, existuje i solubilní forma (extracelulární doména- v cirkulaci- vazebný protein pro leptin?) • Jeho chybění vede k morbidní obezitě • Hladiny leptinu pozitivně korelují s obsahem tuku v organizmu (↓štíhlí, obézní)-informace o stavu tukových zásob • Regulátor příjmu potravy a energetického výdeje • Periferní účinky - ovlivnění inzulinové senzitivity
Regulace příjmu potravy Neuropeptid Y Melanocyty stimulující h.
Leptin a obezita • Deficit leptinu není epidemiologicky signifikatní příčinou obezity (3 popsané případy mutací leptinového genu u člověka spojené s morbidní obezitou) • U většiny obézních pacientů je přítomna hyperleptinémie tj. hladiny leptinu korelují s obsahem tuku v organizmu • Redukce hmotnosti vede také k poklesu hladin leptinu • Studie s podáváním leptinu obézním subjektům s hyperleptinémií nepotvrdily jednoznačný přínos při redukci hmotnosti
Proč hyperleptinémie netlumí u obézních nemocných příjem potravy: • Důvodem je leptinorezistence buď na úrovni transportu leptinu přes hematencefalickou bariéru nebo na postreceptorové úrovni • Leptin není fylogeneticky primárně určen k tlumení příjmu potravy ale k adaptaci organizmu na dlouhodobé hladovění
Mutace leptinového genu u lidí vede k morbidní obezitě, substituce leptinu fenotyp upravuje Farooqi SI, J. Clin. Invest.110:1093-1103 (2002)
ob/ob myši • Nepřítomnost leptinu v organizmu způsobuje morbidní obezitu spojenou s inzulínovou rezistencí. • Obě tyto myši stejného věku mají defektní ob gen kódující leptin. • Myši na obrázku vpravo byl injekčně podáván purifikovaný leptin a její hmotnost dosáhla 35 gramů. • Myš na obrázku vlevo bez leptinové substituce vážila 67 gramů a byla také méně fyzicky aktivní
Potkani kmene Koletsky • Obezita vyvolaná mutací leptinového receptoru u potkana kmene Koletsky • Mutace způsobuje vymizení extracelulární domény tohoto receptoru (na obrázku vpravo) • Vlevo normální potkan kmene Wistar stejného věku
cílovými proteiny G-proteinových podjednotek jsou: • iontové kanály • enzymy vázané na membránu
nejčastějšími cílovými enzymy G-proteinů jsou: • adenylátcykláza tvorba cAMP • fosfolipáza C tvorba inositoltrifosfátu a diacylglycerolu
Adenylyl(lát) cykláza = přeměňuje ATP na cAMP = cyklický AMP = adenosin-3´,5´-cyklický monofosfát
Ras protein • Původ názvu: RAt Sarcoma • Kódován stejnojmenným genem, protoonkogen (Mu ve 20-30% lidských malignit) • Zřejmě přítomen u všech eukaryot • Stimuluje proliferaci či diferenciaci B
Aktivace proteinu Ras aktivovanou receptorovou tyrosinkinázou
Langerhansovy ostrůvky (insulae pancreaticae) • Morfologicky i funkčně odlišné oblasti pankreatu • Přítomny v endokrinní části pankreatu. • V dospělosti okolo 1 mil. • Složen z několika stovek polygonálních buněk oddělených od sebe četnými kapilárami • 3 základní typy buněk: • α-buňky A • β-buňky B • δ-buňky D Langerhansův ostrůvek kolem exokrinní buňky pankreatu
Inzulín • Z lat. Insula - ostrov • Hormon produkovaný β-buňkami Langerhansových ostrůvků slinivky břišní • Odtud se vylučuje do krve • Váže na inzulínové receptory, které jsou uložené zejména na povrchu jaterních, svalových, nebo tukových buněk. • Některé buňky v těle přijímají glukózu i bez přítomnosti inzulínu, například nervové buňky. • Snižuje hladinu glukózy v krvi (glykémii) • V játrech naopak podporuje syntézu jaterního glykogenu. β-buňky
Vznik inzulínu 1. fáze: vzniká pre-proinzulin na ribozomech Langerhansových ostrůvků 2. fáze: v endoplazmatickém retikulu je pre-proinzulin přeměňován na proinzulin, který je tvořen řetězci A a B spojenými C-peptidem (connection peptid) 3. fáze: proinzulin putuje do sekrečních granul β-buněk, kde je v Golgiho aparátu rozštěpen na C-peptid a inzulín 4. fáze: inzulín skladován do času potřeby v sekrečních granulích β-buněk Je tvořen 51 AMK
Autoimunitní destrukce -buněk Langerhansových ostrůvků pankreatu