Download

1 / 17
170 likes | 253 Views
Apoptosis. Many functions for programmed cell death, many consequences for failure of its regulation Apoptosis vs. necrosis Intrinsic and extrinsic pathways for survival vs. proliferation vs. apoptosis Caspases: activation by proteolysis Mitochondria and cytochrome c release BCL-2 proteins.

E N D
Apoptosis • Many functions for programmed cell death, many consequences for failure of its regulation • Apoptosis vs. necrosis • Intrinsic and extrinsic pathways for survival vs. proliferation vs. apoptosis • Caspases: activation by proteolysis • Mitochondria and cytochrome c release • BCL-2 proteins Jason Kahn: Apoptosis
What is Apoptosis? • Greek: Falling away • A.k.a. Programmed Cell Death or PCD • Distinct from Necrosis, which is unplanned cell death • Apoptosis is actually the default pathway for metazoan cells: they must constantly be told to keep living • Sources: • Voet and Voet Chapter 34 • Helmreich Chapter 13 • Gomperts Chapter 14 • Danial and Korsmeyer (2004), “Cell Death: Critical Control Points.” Cell116, 205–219. • Jiang and Wang (2004), “Cytochrome C-Mediated Apoptosis.” Annu. Rev. Biochem.73, 87-106. • http://www.roche-applied-science.com/sis/apoptosis/ From http://www.roche-applied-science.com/sis/apoptosis/ Jason Kahn: Apoptosis
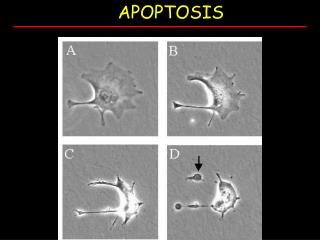
Apoptosis vs. Necrosis • Necrotic cells lose their ability to handle osmotic pressure, swell, and burst, much like cell lysis done in the lab. Their contents spill into the surroundings, tending to produce inflammation. DNA is largely intact, possibly capable of being taken up by neighboring cells (…bad). • Apoptosis is characterized by contraction of the cell and “blebbing” (blistering) of membrane patches, DNA degradation, mitochondrial disruption and fission, and eventual disintegration into encapsulated blobs of stuff. These capsules (apoptotic bodies) are readily absorbed by macrophages or other neighboring cells withoutleading to inflammation. • Apoptosis is an orderlyprocess, often part of normal development: many pathways werefirst worked out in C. elegans,in which 131 out of 1090cells die during thedevelopment of the matureworm. Jason Kahn: Apoptosis
Progression of Apoptosis • Cell is committed to apoptosis upon cytochrome c release, even though it takes a couple of hours to complete the process • This is not a process that has been selected to be one that can be aborted half way through! • Hence we will seerather aggressivesignalling, suchas proteolysis,membrane pores • Compare to bloodclotting, cell cycleprogression: alsounidirectional. Jason Kahn: Apoptosis
Functions for Apoptosis • Development: Normal development requires transient tissues. For example, mammalian embryos have webbed feet. Self-reactive immature T cells are eliminated. Neurons that have not made enough synapses are eliminated. • Apoptosis is part of the body’s defenses against viral invasion: infected cells can be induced to apoptose by T cells that recognize displayed antigens, excessive DNA damage leads to apoptosis. The mouse on the right lacks Caspase 9 V+V 3/e, Chapter 34 Jason Kahn: Apoptosis
Removal of Apoptotic Bodies • Exposure of phosphatidyl serine is the “eat me” signal Jason Kahn: Apoptosis
Proliferation vs. apoptosis • Typically the number of cells that divide each day is equal to the number that die (steady state). • Thus there is a delicate balance between proliferation and apoptosis, and many signalling pathways (e.g. c-myc) activate both, so that rapidly dividing cells are often short-lived. Jason Kahn: Apoptosis
What Can Go Wrong? • Excessive apoptosis is seen in autoimmune diseases like Type I diabetes, in neurodegenerative disorders like Parkinson’s and Alzheimer’s, and in infertility and immunodeficiency • Insufficient apoptosis can lead to autoimmunity and to cancer: apoptosis is supposed to be the last resort for cells that have lost the ability to respond to their surroundings, but if the apoptotic pathways themselves break down… For example, BCL-2 = B-cell lymphoma 2 is an antiapoptotic protein. It is overexpressed after a common chromosomal translocation event, and the resulting cells are resistant to apoptosis and hence prone to uncontrolled proliferation if other mutations occur that dysregulate cell division. • Viruses have strategies to block apoptotic signalling • Cancer cells are often found to overexpress “IAP” proteins = inhibitors of apoptosis. Jason Kahn: Apoptosis
What Can Go Wrong II Tumor Necrosis Factor family members control apoptosis, proliferation, and other cascades Aggarwal (2003). “Signalling pathways of the TNF superfamily: a double-edged sword.” Nature Reviews in Immunology, 3 , 745 Jason Kahn: Apoptosis
Apoptosis Is Complex (Roche web site) Jason Kahn: Apoptosis
Intrinsic vs. Extrinsic Pathways • Extrinsic: signals from outside the cell can lead to survival, proliferation, or apoptosis • Many tissue culture cells die unless they are attached to a surface (the extracellular matrix or ECM). The antiapoptotic signals are mediated through integrins associated with focal adhesion sites. • Cytotoxic T cells express ligands (e.g. the Fas ligand) which activate “death receptors” on the surface of target cells to induce apoptosis. • Intrinsic: signals from inside the cell, or lack of survival signals from outside • When DNA is damaged, activation of p53 leads to increased DNA repair and cell cycle arrest, but higher levels of damage activate apoptosis. • This is also the default pathway for apoptosis in the absence of the growth factors, cytokines, or contacts to the ECM that are needed to keep the cell alive. • Intrinsic and extrinsic pathways are mediated through the mitochondria: cytochrome c is released. • Both types of pathway end up activating Caspases (Cysteine-ASPartate-Proteases), which are cysteine proteases that cut after DEVD sequences. Jason Kahn: Apoptosis
Caspases • Very specific proteases. They are synthesized as zymogens (procaspases), activated by proteolytic cleavage. They can then activate other caspases or cleave other targets. • Initiator caspases (-8, -9, -10) can self-activate slowly. They have effector caspases as their targets. Confers amplification and irreversibility. • Effector caspases (-3, -6, -7) cut up many other targets (~60), including: • Inhibitors of apoptosis • The ICAD inhibitor of the CAD DNase, which destroyschromosomal DNA (easy assay ->) • BCL-2, an antiapoptotic protein • Cytoskeletal proteins and the nuclear lamin • DNA replication and repair proteins like RFC (theeukaryotic clamp loader) • Repair enzymes and splicing factors • Transcription factors and kinases DED = death effector domain Jason Kahn: Apoptosis
Extrinsic Pathways Trimerization of receptors upon ligand binding leads to trimerization of cytoplasmic death domains, which bind adaptors, which recruit caspase 8 through its DED domains TNF is the founding extracellular ligand, acting through the TNFR1 receptor Jason Kahn: Apoptosis
Intrinsic Pathway • Release of cytochrome c from the mitochondria is the signal: it’s never foundoutside otherwise. It assembles with the adapter APAF-1 to make the “apoptosome,” which activates Caspase-9, which then cleaves Caspase 3. • The antiapoptotic BCL-2 blocks Cytochrome c release. Jason Kahn: Apoptosis
APAF-1/Cytochrome C + Caspase-9 Figure 34-117b Cryoelectron microscopy–based images of the apoptosome at 27-Å resolution. (b) The apoptosome in complex with a noncleavable mutant of procaspase-9 in oblique top view. Jason Kahn: Apoptosis
Mechanism of Cytochrome C Release • Mechanism of release is controversial. There is a family of BCL-2 related proteins. Group I: antiapoptotic, Groups 2 + 3: proapoptotic. • Some BCL-2 related proteins form pores in membranes. But are they big enough? And why would antiapoptotic factors act as pores? Bax/Bak may aggregate to form a large pore. • Interaction between Bax and/or Bak and VDAC = voltage-dependent anion channel might form pores? • Simple dissipation of protein gradient or introduction of other molecules? • Pro-apoptotic proteins may also act by sequestering BCL-2 or BCL-XL: there are several proteins that can interact with each other through their BH3 domains: Jason Kahn: Apoptosis
More on BCL-2 • BCL-2 sequesters APAF-1 either directly or by preventing cytochrome c release, thereby keeping the procaspases bound to APAF-1 inactive. • Inhibitors of BCL-2 (such as Bad), either compete with APAF-1 for binding to BCL-2 or promote release of cytochrome c, leading to APAF-1 activation via cytochrome c binding, multimerization of APAF-1, and caspase activation Jason Kahn: Apoptosis