Download
1 / 31
310 likes | 411 Views
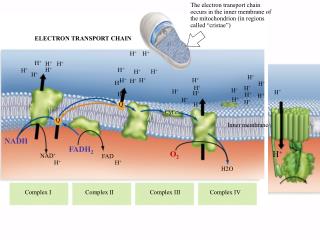
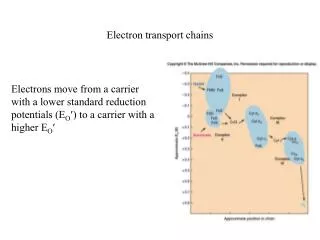
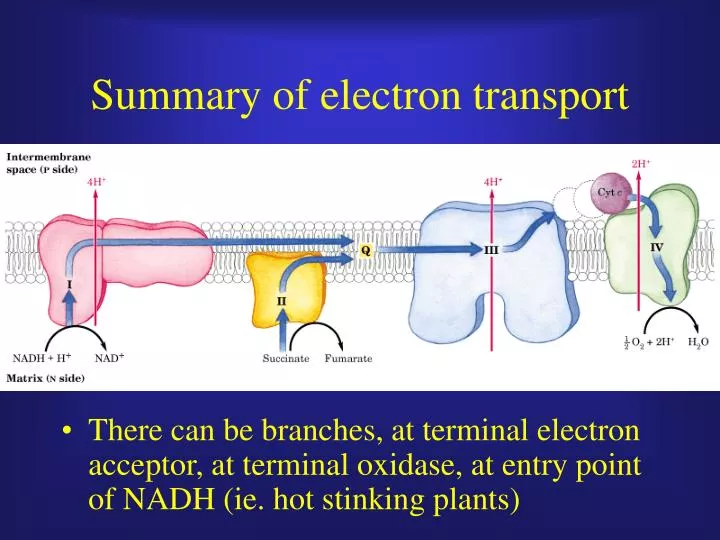
Summary of electron transport. There can be branches, at terminal electron acceptor, at terminal oxidase, at entry point of NADH (ie. hot stinking plants). Unfortunately, oxygen is not just a terminal electron acceptor. NADH, a great source of energy.

E N D
Summary of electron transport • There can be branches, at terminal electron acceptor, at terminal oxidase, at entry point of NADH (ie. hot stinking plants)
Unfortunately, oxygen is not just a terminal electron acceptor
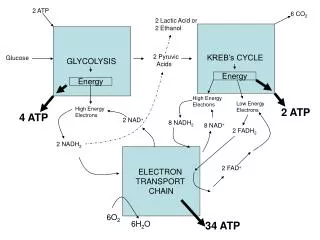
NADH, a great source of energy • NADH + 11 H+ + ½ O2 NAD+ + 10 H+ + H2O • Highly exergonic; DGo = -220 kJ/mol • Actually in cell, much NADH than NAD, making the available free energy more negative • Much of this energy is used to pump protons out of the matrix
Pumping protons lowers the pH and generates an electrical potential
Generation of a proton-motive force • In an actively respiring mitochondria, the pH is ~0.75 units lower outside than in the matrix • Also generates an electrical potential of 0.15 V across the membrane, because of the net movement of positively charged protons outward across the membrane (separation of charge of a proton without a counterion) • The pH difference and electrical potential both contribute to a proton motive force
Really, what does that mean? • Energy from electron transport drives an active transport system, which pumps protons across a membrane. This action generates an electrochemical gradient through charge separation, and results in a lower pH outside rather than in. Protons have a tendency to flow back in to equalize the pH and charge. This flow is coupled to ATP synthesis.
Measuring the proton motive force DmH = Dy – 2.3RTDpH/F (different in Lehninger) mH is the resulting proton motive force (sometimes p) y is the electrochemical membrane potential pH has a negative value, thus contribution is positive in this equation
Don’t get bogged down in the math, but … • (under standard conditions) DmH = 0.224 V Plug into DGo = -nFDEo and DGo = ~20 kJ/mole H+ The bottomline
The proton motive force bottom line Two components to energy derived from electron transport, pH and electrical potential. The electrical potential is the primary contributor to free energy. Most of the energy from oxidation of NADH is conserved in the proton gradient
Electron transfer and ATP synthesis are coupled • ATP synthesis occurs only if electron transfer does, and vice-versa • When isolated mitochondria are suspended in buffer containing ADP, Pi and an oxidizable substrate (succinate) three things happen • Substrate is oxidized • Oxygen is consumed • ATP is synthesized
All components are essential • If ADP were omitted, no ATP synthesis would occur and electron transfer to oxygen does not proceed, as well. Black – oxygen consumption Red – ATP synthesis
There are compounds that can inhibit ATP synthesis • The antibiotic oligomycin binds to ATP synthase and inhibit it’s action. • By stopping ATP synthesis, this compound also stops electron transport. • Because oligomycin is specific for ATP synthase and not the various electron carriers, this inhibition supports the coupling of ATP synthesis to electron transport
There are compounds that can uncouple ATP synthesis from electron transport • DNP and FCCP block ATP synthesis, while permitting continued electron transport to oxygen – they are uncouplers • They pick up protons from the outside, diffuse in (they are hydrophobic so can pass through the membrane), and release proton back inside. • Electrons are still passed through the electron transport chain, but the proton gradient is destroyed.
Something we’ll cover when we talk about enzymes in detail: • ATP synthase stabilizes ATP relative to ADP + Pi by binding ATP more tightly, this results in a free energy change that is near zero • This is an important point, but ignore for the most part now as we will cover this in detail later • What’s important now is that this reaction ATP synthesis from ADP and Pi occurs without a huge input of energy – you’ll see it is just mechanical energy.
ATP synthase has two functional domains • This enzyme has two distinct parts, one a peripheral membrane protein (F1) and one a integral membrane protein (Fo) ( the o stands for oligomycin sensitive) • These parts can be separated biochemically, and isolated F1 catalyses ATP hydrolysis (it has the site for ATP synthesis and hydrolysis)
The F1 component • This component is made up of nine proteins of five different types with a composition of: a3b3gde • Each of the three b subunits have a catalytic or “active” site where the reaction occurs • ADP + Pi ATP + H2O
The a and b subunits make a cylinder with the g subunit as an internal shaft
Conformational changes • Although the b subunits have the exact same amino acid sequence and composition, they are in different conformations due to the g subunit. • These conformational differences affect how the enzyme binds ATP and ADP
Rotation of the g subunit by H+ translocation drives ATP synthesis • Passage of protons through the Fo component causes g to rotate in that internal chamber • Each rotation of 120o causes g to contact another b subunit, this contact forces b to drop ATP and stay empty • The three b subunits interact so that when one is empty, one has ADP and Pi, while another has ATP.
Proton transfer is converted to mechanical energy, then chemical energy
ATP synthase – at work • http://nature.berkeley.edu/~hongwang/Project/ATP_synthase/ • http://www.sciencemag.org/feature/data/1045705.shl
ATP exits the mitochondria through active transport • P N Side Side
Regulation of ETC • Rate of mitochondrial respiration controlled by ADP availability ([ATP]/[ADP][Pi]) • IF1 can bind and block ATP synthase at low pH • Hypoxia influences gene expression
Coordinated regulation – more on this later, but think about global effects
What happens when… • Cells increase NADH oxidation using alternative NADH oxidase? • Cells using lots of ATP