Download

1 / 43
600 likes | 1.87k Views
Enzim Aktivitesinin Regülasyonu. Dr. Gülçin Eskandari. Canlı varlıklar kendi metabolik yolaklarını kontrol etme yeteneğine sahip Bu kontrol mekanizmasını bazı spesifik enzimlerin konsantrasyonlarını ve aktivitelerini düzenleyerek yaparlar

E N D
Enzim Aktivitesinin Regülasyonu Dr. Gülçin Eskandari
Canlı varlıklar kendi metabolik yolaklarını kontrol etme yeteneğine sahip • Bu kontrol mekanizmasını bazı spesifik enzimlerin konsantrasyonlarını ve aktivitelerini düzenleyerek yaparlar • Enzimlerin çoğu protein yapıda olduklarından, enzim konsantrasyonları protein sentez ve degredasyonu arasındaki dengeye bağlı
Spesifik proteinlerin sentezi mRNA sentezi ve mRNA’nın translasyonda kullanımını da kapsayan birçok basamakta kontrol edilir • Protein degredasyon hızı ise protein degredasyonundan sorumlu enzimlerin konsantrasyon ve aktiviteleri gibi mekanizmalarla kontrol edilir
Protein sentez ve degredasyon kontrolü ile olan mekanizmalar uzun dönemde etkili olmakta • Bazı durumlarda çok çabuk değişen koşullara organizma yanıt vermekte • Hızlı yanıtın oluşması regulatuvar enzimler üzerindeki metabolik kontrolle gerçekleşir • Birçok enzim aktivitesi pH değişikliğinden etkilenir • ÖR: Lizozomal enzimler pH 5’te en yüksek aktiviteyi gösterirlerken, intrasellüler pH olan 7’de aktiviteleri oldukça azalır • Ortam pH değişikliği de pratik bir yol değil
Regulatuvar Enzimler • Metabolizmada ardışık yollar bulunmakta ve birçok enzim bu yollarda görev almakta • Birinci enzimin katalizlediği reaksiyonun ürünü, bir sonraki basamakta görev alan enzim için substrat olmakta • Bu ardışık reaksiyonlarda hız kısıtlayıcı basamağı oluşturan ve en yavaş katalizin olduğu bir basamak bulunur • Bu tip basamaklarda görev alan enzimlere regulatuvar enzimler denir
Regulatuvar Enzimler • Regulatuvar enzimler gelen sinyallere göre katalitik aktivitelerini artırır ya da azaltırlar • Multienzim sistemlerinin çoğunda ilk reaksiyonu katalizleyen enzim bir regulatuvar enzim • Bu ardışık sırada yer alan diğer enzimler, kendilerine substrat geldiği oranda aktivite gösterirler
Regulatuvar Enzimler • Regulatuvar enzimlerin aktiviteleri değişik tipte sinyal molekülleri ile düzenlenir • Bunlar genellikle küçük metabolitler ya da kofaktörler • Metabolik yolaklarda enzim aktivitelerinin düzenlenmesinde iki strateji izlenmekte; • Enzimin kovalent yapısında değişiklik oluşturularak Km ya da kcat değerlerinde değişiklik oluşturulması • Effektör olarak adlandırılan inhibitör ya da aktivatör bir molekülün enzime reversible olarak bağlanması ile Km ya da kcat değerlerinde değişiklik oluşturulması
Parsiyel Proteoliz: İrreversible bir kovalent modifikasyon • Pankreas tripsin, kimotripsin, elastaz ve karboksipeptidazları inaktif zimojenler olarak sekrete eder • Bunlar ekstrasellüler olarak diğer proteazlar tarafından parsiyel proteolize uğratıldıklarında aktivite kazanırlar • Tripsin, intestinal bir enzim olan enterokinazın N terminal hekzapeptidi yapısından ayırmasıyla aktive kazanır • Tripsin ise kimotripsinin N terminal ucundan 15 amino asidlik bir diziyi kopararak onu aktif hale getirir
Parsiyel Proteoliz • Sindirim enzimlerinin bu şekilde geç aktivite kazanmalarının faydası, sentezlendikleri doku olan pankreasa zarar verememeleri • Substrat bağlanma cebi zimojen formda tam olarak uygun olmadığından, zimojen formun Km değeri daha yüksek
Parsiyel Proteoliz • Pıhtılaşma kaskadında fonksiyon gören enzimler de parsiyel proteolizle aktivasyon kazanırlar • Bunun önemi kaskada gereksinim anına kadar inaktif formda kalmaları • Pıhtılaşama kaskadında fonksiyon gören serin proteaz enzimleri bulunmakta
Parsiyel Proteoliz • Enzimlerin parsiyel proteolizle aktivasyon kazanmaları irreversible bir olay • Enzim aktivasyon kazandıktan sonra degrade ya da inhibe edilene kadar aktif • Bu özellikle pıhtılaşma kaskadında pıhtılaşmanın etraf dokulara da yayılması açısından önem taşıyabilir
Parsiyel Proteoliz • Ancak birçok mekanizma pıhtılaşmayı durdurmak için çalışır • Aktif enzim konsantrasyonlarının kan akımı ile dilue edilmesi • Aktif enzimlerin kan akımıyla karaciğere taşınarak degrade edilmeleri • Pıhtılaşmayı engellemekle görevli diğer kan proteinlerinin (protein C ve S gibi) aktif enzimlerin aktif bölgelerine bağlanarak inhibe etmeleri
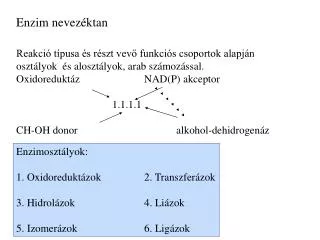
Fosforilasyon • Enzimlerin serin, treonin ya da tirozin residuelarından fosforilasyonu oldukça sık görülen bir kovalent modifikasyon türü • Fosforilasyonu spesifik protein kinazlar, defosforilasyonu ise fosfatazlar geçekleştirir • Kinazlar ve fosfatazların kendileri de metabolik regülasyon altında
Fosforilasyon • Bu enzimler hormonlar ya da büyüme faktörleri gibi ekstrasellüler sinyallerle fosforile ya da defosforile edilirler • ÖR: Epinefrin kas ve adipoz dokuda cAMP-bağımlı protein kinazlar üzerinden bazı enzimlerin fosforilasyonla aktivasyonuna ya da inaktivasyonuna neden olur • Bazı enzimler fosforilasyonla aktifleşirken, bazıları da fosforilasyonla inaktifleşirler
Epinefrin • Adipoz dokuda TAG Lipaz TAG Lipaz-P (aktif) FFA • Kas dokusunda Glikojen Fosforilaz Glikojen Fosforilaz-P (aktif) Glukoz Glikojen Sentaz Glikojen Sentaz-P (inaktif)
Fosforilasyon • Bir enzimin fosforilasyonla allosterik effektörlere olan duyarlılığı artabilir ya da azalabilir • Glikojen sentazın fosforilasyonu ile enzimin glukoz-6-P ve Pi’la inhibisyona olan duyarlılığı artar • Glikojen fosforilazın fosforilasyonu enzimin allosterik aktivatörü olan AMP’ye duyarlılığını azaltır
Adenilasyon • Enzimin tirozin residuesuna adenil (AMP) grubunun eklenmesi bir diğer reversible kovalent modifikasyon türü • Büyük negatif yüklü bir adenil grubunun eklenmesi enzimi inhibe eder • Belki de bu etkisini enzimin aktif bölgesini kapatarak gerçekleştirir
Kovalent Modifikasyon “Kovalent modifikasyon bir hücrenin metabolik yolaklarında görev alan enzimler üzerinde hızlı regülasyon yapmasına olanak tanır”
Allosterik Regülasyon • Ekstrasellüler sinyallere yanıt olarak genellikle fosforilasyon kullanılırken, hücre içi değişen koşullara yanıt olarak genellikle allosterik regülasyon kullanılır • ATP, bir amino asid ya da metabolik bir ara ürünün fazlalığı ya da azlığı allosterik regülasyonu başlatabilir • Kovalent modifikasyondan farklı olarak, herhangi bir enzim etkisine gereksinim duymaz
Allosterik Regülasyon • Metabolik yolaktaki bir ara ürün allosterik effektör olarak fonksiyon görüyor ise, enzim aktivitesi effektör konsantrasyonu ile düzenlenir • Allosterik regülasyon metabolizmada ince ayar yapar
Allosterik Enzimler • Allosterik enzimlerde, modülatör (effektör) adı verilen regulatuvar metabolitler enzime reversible, nonkovalent olarak bağlanır • Modülatörün enzime bağlanması, enzimde konformasyonel değişikliğe neden olur • Konformasyonel değişiklik de ya aktivitenin artması ya da azalmasıyla sonuçlanır
Allosterik Enzimler • Allosterik enzim modülatörleri inhibitör ya da stimülatör olabilirler • Stimülatör genelde substratın kendisi • Substratı ve modülatörü aynı olan regülatuar enzimlere homotropik denir Farklı olanlara ise heterotropik denir • Bazı enzimlerin ise iki ya da daha fazla sayıda modülatörü olabilir • ATP, ADP, AMP ve Pi birçok anahtar enzimin regülatörü • Bu moleküller enzimin aktif bölgesi dışında allosterik bölgesine bağlanarak etki ederler
AllosterikEnzimler • Allosterik enzimler özellik olarak basit enzimlerden farklılık gösterirler • Bunlar yapısal farklılıklar da olabilir • Allosterik enzimlerin aktif ya da katalitik bölgelerine ek olarak, modülatör bağlanma bölgeleri de bulunur • Bu bölgeye allosterik bölge denir • Substrat bağlanma özgünlüğü gibi, bu bölgenin de modülatör bağlama özgünlüğü bulunur • Birden çok modülatörü olan enzimlerin her bir modülatörü için ayrı bağlama bölgesi var • Homotropik enzimlerde ise aktif ve modülatör bölge aynı
Allosterik Enzimler • Allosterik regülasyonla aktiviteleri düzenlenen birçok enzimin birden fazla subuniti var • Birçok durumda enzim aktivitesindeki değişiklik subunitler arası etkileşimle ilgili
Kooperatif Bağlanma • Bunun için öne sürülen iki model bulunmakta • Simetri modelinde ya tüm subunitler aktif ya da hepsi inaktif • Ardışık modelde ise subunitler tek tek de konformasyonel değişiklik gösterip aktif hale geçebilmekte Tek bir substratın bağlanması da diğerlerinde konformasyonel değişikliği aktive etmekte ve diğer bölgelere substrat bağlanması kolaylaşmakta
Ardışık Model
Allosterik Enzimler • Allosterik enzimler kinetik açıdan da normal Michealis-Menten davranışından farklılık gösterir • Hiperbolik yerine sigmoidal satürasyon eğrileri var • Oksijenin hemoglobine kooperatif bağlanmasına benzer • Bu eğride Vmax/2’ye karşılık gelen substrat konsantrasyonu Km olarak adlandırılamaz • Bunun için K0.5 terimi kullanılır
Allosterik Enzimler • Sigmoid kinetik davranışı, multiple protein subunitlerine kooperatif bağlanma özelliği gösterir • Diğer bir deyişle bir subunitteki yapısal değişiklik, komşu subunitlerdeki değişikliğe neden olmakta • Hemoglobine oksijen bağlanmasında gözlenen olaylar gibi • Myoglobinin hiperbolik saturasyon eğrisi vermekte • Oysa hemoglobinin 4 oksijen bağlama bölgesi olup, allosterik enzimler gibi sigmoidal saturasyon eğrisi göstermekte
Fosfofruktokinaz • Enzim dört subunitten oluşur • AMP enzimin allosterik effektörü • Ortamda ATP varlığında substrat olan fruktoz-6-P konsantrasyonuna bağlı olarak sigmoidal kinetik davranış sergiler • Çok düşük ATP varlığında ya da allosterik effektörü olan AMP varlığında hiperbolik kinetik gösterir • Allosterik effektörler kinetik değişikliğine neden olabilirler
İki subuniti bulunan allosterik bir enzime substrat bağlanması
Allosterik Feed-Back İnhibisyon • Bazı multienzim sistemlerinde regulatuvar enzimler, yolağın sonunda oluşan son ürün ile spesifik olarak inhibe edilir • Son ürün oluşumu hücre ihtiyacını aştığı her durumda feed-back inhibisyon oluşabilir • Bu olayda regulatuvar enzimin katalizlediği reaksiyon yavaşlar ve bu aşamadan sonra görev alan tüm enzimler de düşük aktivite gösterir • Böylece yolağın son ürünü hücrenin ihtiyacı düzeyinde tutulur • Bu tür düzenlemeye de allosterik feed-back inhibisyon denir