Download

1 / 38
420 likes | 638 Views
Chapter 10 Dynamics of Adaptive Immunity. This chapter tries to integrate much of what we have already covered so we can better understand how the immune system protects from infection. How infectious agents cause disease The course of an adaptive immune response Immunological memory.

E N D
Chapter 10 Dynamics of Adaptive Immunity This chapter tries to integrate much of what we have already covered so we can better understand how the immune system protects from infection. • How infectious agents cause disease • The course of an adaptive immune response • Immunological memory
The course of a typical infection and adaptive immune responses can be divided into phases If no innate immunity
The establishment of an infection depends on several factors: • Characteristics of the microorganism • Number of organisms • Mode of transmission (how and where they contact the host) • Stability of the organism (in and outside of the host)
Almost all infections begin at an epithelial surface* Skin and tight junctions usually prevent penetration *skin or mucosal surface of the respiratory, gastrointestinal, and urogenital tract which are always exposed to microorganisms.Interstitial tissues are normally free of any microorganisms (sterile)
Most pathogens have developed defenses to partially escape destruction by the innate protective mechanisms listed in boxes 1 & 2 below Most pathogens cause disease in only one or a few related species because the pathogens can adhere to epithelial surfaces of only a few species or can escape innate immunity of only a few species.
The course of the adaptive response to infection How do infections progress in immune deficient individuals? Adaptive immune response becomes important when innate immunity cannot deal with the infection within a couple of day
The results of an immune response vary depending on the individual and the pathogen and other factors (e.g, level of care). • After a primary immune response, usually there are no pathogens remaining. • Sometimes the infection is contained but not elimintated (TB granuloma, herpes virus becomes latent, HIV mutates). • Sometimes the pathogen or the immune response, or both, leave significant tissue damage (e.g., scaring, blindness). • Often life-long immunity
Non specific response of innate immunity for adaptive immunity (bacterial infection) macrophage activation Ag uptake by DC complement activation DC activation chemokine, cytokine MHC class II ↑ B7-1,B7-2 expression cell migration and other response Peripheral lymphoid tissues Inflammation Initiation of adaptive immunity T cell activation effector T cell infection sites CCL19,20,21 in
Cytokines made in the later stages of an infection influence differentiation of CD4 T cells toward TH1 or TH2 cells - The differentiation of naive Th cells into the two major classes of CD4 effector cells can be profoundly influenced by cytokines present during the initial proliferative phase of T cell activation - CD4 T cell + IL-12 + IFNg ⇒ TH1 - Signals that stimulate dendritic cells to release IL-12 include CCL3 (MIP-1a),CCL4(MIP-1b),CC5(RANTES) - Production of IL-12 by dendritic cells is also stimulated by IFNg, Prostaglandin E2, LPS (TLR-4) DC, macrophage NK ,CD8 T cell ↓ CCR5, CCR1 on DC ↑ DC,macrophage, endotherial cells
NK1.1 T cell is a T cell sub-set which expresses a surface molecule usually associated with NK cells and do not express normal TCR repertoire NK cell TH1 TH2 What governs whether a TH0 cell differentiates into TH1 or TH2? Pathogens influence cytokines that affect TH0 differentiation into TH1 or TH2
- Mice deficient in MyD88 (TLR signaling component) do not survive a challenge with Toxoplasma gondii (survive inside macrophage vesicle and induce TH1 response) ⇒ due to failure in IL-12 production - CD4 T cell + IL-4 +(IL-6) ⇒ TH2⇒ IL-4 mast cell, NKT cell , DC+TLR-2 ligand, Notch signaling
The distinct subset of CD4 T cell can regulates each other’s regulation - When IFN-g or IL-4 are present, TGFb and IL-6 do not efficiently generate TH17 cells - IL-10 from TH2 inhibition of TH1 development by suppressing IL-12 production by DC - IFNg from TH1 inhibition of TH2 development - Certain responses are dominated by either a TH2 or TH1 response. However, under many circumstance in vivo there is a mixed TH1 and TH2 response - NK T cells might regulate TH1 vs TH2 development - NK IFNg TH1 response IL-4 → TH2 response IFNg APC NKT → CD1 Ag:glycolipid mycolic acid • CD4+ • CD4-CD8-
T cell subsets produce cytokines that regulate development of other subsets For example, TH1 inhibits TH2 and TH2 inhibits TH1. Thus most responses are dominated by either TH1 or TH2 and are not balanced responses
Leishmania Leishmania anti-IL-4 Ab - The importance of TH1, vs TH2 response in mice BALB/C mice TH2 response → die C57BL/6 mice TH1 response → survive BALB/C mice TH1 response → survive - CD8 T cells can also respond to antigen by secreting cytokines typical of either TH1 or TH2 ⇒ Tc1 or Tc2 In leprosy infection , tuberculoid leprosy : Tc1 → induction of TH1 → macrophage activation → not serious lepromatous leprosy : Tc2 → IL10,TGFb → suppression of TH1→ severe disease - The nature and amount of antigenic peptide can also effect the differentiation of CD4 T cells large amount of peptide → TH1 low amount of peptide → TH2 (eg. Allergen)
Cytokines influence the CD4 T cell subset development and this can be crucial for development of an appropriate immune response Mice normally respond inappropriately to Leishmania and thus are killed by the parasite
Migration of effector T cells to site of infection - Most of the antigen-specific effector T cells cease production of L-selectin and express VLA-4 which binds to VCAM-1 that is induced on activated endothelial cells ⇒ initiation of the extravasation - In the absecense of a previous inflammatory response, activated T cell enter all tissues via P-selectin : PSGL-1 interaction - Effector T cells that recognize pathogen antigens in the tissue → production of cytokines (eg TNFa) → activation of endothelial cell → E-selection,VCAM-1, ICAM-1,CCL5 expression → Action on effector T cells → activation of adhesion molecules on T cells - TNFa and IFNg released by activated T cells → act synergistically to change the shape of endothelia cells
Effector T cells change their adhesion molecules so they can migrate to the site of an infection. That is, adhesion molecules regulate trafficking
- Differential expression of adhesion molecules can direct different subsets of effector T cells to specific sites : The selective expression of different homing receptors that bind to tissue-specific addressins • Some activated T cells express cutaneous lymphocyte antigen (CLA) ⇒ binds to E-selectin on cutaneous vascular endothelium Figure 10.10 • CLA expressing T lymphocytes also produce CCR4 ⇒ binds CCL17(TARC) present at high levels on the endotherium of cutaneous blood vessels • Skin-homing T cells carry CCR10 (GPR-2) ⇒ binds CCL27 on keratinocytes
CD4+ T cells can license dendritic cells to activate CD8+ T cells (CTLs) (figure 8.28) but some CD8+ cells can be activated by dendritic cell in the absence of CD4+ T cells CD8 T cells can be induced to secrete INF-g in an antigen non-specific way
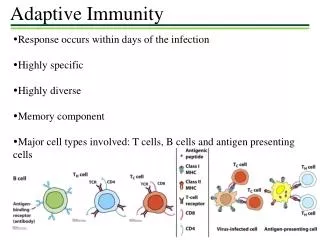
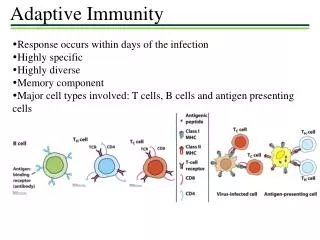
Antibody responses under the direction of helper T cells - B cells specific for a protein antigen cannot be activated to proliferate, form germinal centers or differentiate into plasma cells until they encounter a helper T cell that is specific for one of the peptide derived from that Ag - The specialized regions of peripheral lymphoid tissue provide an environment where antigen specific naïve B cell can interact with helper T cells Figure 10.14 - Antibodies secreted by B cells early in the response are important in trapping antigen in the form of Ag • Ab complex ⇒ become coated with complements ⇒ binds to Fc, complement receptors on FDC ⇒ stay for very long periods - CXCL13 ↔ CXCR5 ⇒ B cell homing to germinal center CCL19,21 ↔ CCR7 ⇒ B cell interaction with T cell
Peripheral lymphoid organs are the site of antigen-specific B cell activation [antigen binding (signal 1) and T cell help (signal 2)] The book is inconsistent on the location of the primary focus
Antibody responses are sustained in medullary cords and bone marrow • After T-B cell interaction, 90% plasma cell extrafollicular site (2-4day life span) B cell migration to 10% longer life follicular site germinal center formation memory cell plasmablasts bone marrow other peripheral tissue ⇒ plasma cells (months to years life span) ( class switching affinity maturation)
Protective immunity - Different effector mechanisms are used to clear primary infections with different pathogens and to protect against subsequent reinfection ⇒ important in vaccine design Figure 10.16 - protective immunity consists of two components • preformed immune reactants : Abs, effector T cells • long-lived immunological memory - Resolution of an infection is accompanied by the death of most of the effector cells and the generation of memory cells Figure 10.17
Protective immunity is the resistance to a pathogen that results from infection or vaccination. It is due to an adaptive immune response which induced immunological memory of the pathogen. Usually, there are few or no symptoms associated with a infection when it is dealt with by protective immunity Protective immunity consists of preformed immune components and immunological memory
Immunological memory • Immunological memory is long-lived after infection or vaccination - memory reflects a small population of specialized memory cells formed during the adaptive immune response that can persist in the absence of the antigen that originally induced them - the memory response had an approximate half-life of between 8 and 15 years - immunological memory need not be maintained by repeated exposure to infectious virus - Althought most of the memory cells are in a resting state, a small percentage are dividing at any on time cytokines produced either constitutively or during other antigen-specific immune response
Antigen-specific memory B cells differ both quantatively and qualitatively from naïve B cells Figure 10.18 - B cell frequency : 100 fold increase -antibody affinity : higher Figure 10.19 - isotype : Ig G,A,E
No increase in specificity Immunological Memory(maintenance of memory is probably antigen-independent but there is evidence for antigen sequestration or re-exposure to maintain memory) Primary Vs. Secondary B Cell Responses High quality
Primary Vs. Secondary Vs. Tertiary B Cell Responses High quantity High quality
Affinity maturation Affinity maturation is a form of somatic selection (somatic evolution) where B cells, using their surface antibodies (BCRs), compete for limiting amounts of antigen. The B cell must compete with each other and with free antibody produced by plasma cells or plasmablasts. Only those B cells with the highest binding affinity will be able to bind to antigen and thus remain activated.
Some Characteristics of Memory B cells Long-lived Memory B cells have high affinity antibody and high levels of MHC so they are efficient at acquiring antigens (signal 1) and ready to get T help (signal 2) Memory B cells have B7 on their surface so, maybe, they can activate or participate in activation of TH2 (memory) cells Memory B cell have usually switch class from IgM so they usually have other Ig isotypes on their surface (mostly IgG)
Memory T cells remain long after the virus infection is under control* or the virus are gone * The case show here is the reactivation of a letent infection
Naïve and memory T cell have different requirements for survival Naïve T cell require contact with MHC+self peptides for survival (like positive selection in the thymus Antigen encounter (signal 1) plus co-stimulation (signal 2) activates the T cell (proliferation and differentiation) Memory cells can come directly from the activate naïve cells or from the effector cells Some differentiate into effector cells Memory cells need cytokines but not contact with MHC+peptide for survival. This is different from the naïve T cells (top panel) Most effectors die in a few days Memory T cells need MHC+self peptide to proliferate (weak interactions). These remain memory T cells and do not become effectors until they bind the MHC+the correct foreign peptide (strong binding).
Death Plasma cell Naïve B cell Memory B cells Effector T cell Death Naïve T cell Memory T cell
CD4 T cells are required for CTL (CD8) memory but not for primary CTL response
Epitopes ABCD Epitopes ACEF Epitopes ADEG When antigens have both epitopes that were “seen” before and new epitopes, the memory responses to the epitopes seen before will dominate and there may be no response to the new epitopes Original antigenic sin