Download
1 / 1
20 likes | 127 Views
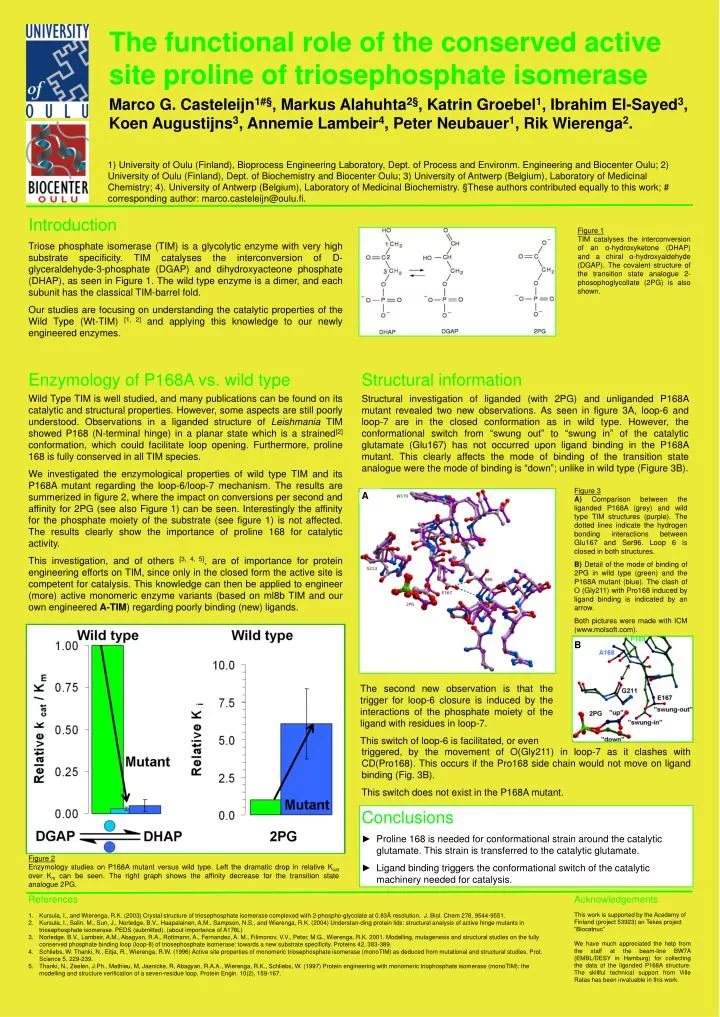
Figure 1. Figure 3. Figure 2. Enzymology studies on P168A mutant versus wild type. Left the dramatic drop in relative K cat over K m can be seen. The right graph shows the affinity decrease for the transition state analogue 2PG.

E N D
Figure 1 Figure 3 Figure 2 Enzymology studies on P168A mutant versus wild type. Left the dramatic drop in relative Kcat over Km can be seen. The right graph shows the affinity decrease for the transition state analogue 2PG. TIM catalyses the interconversion of an α-hydroxyketone (DHAP) and a chiral α-hydroxyaldehyde (DGAP). The covalent structure of the transition state analogue 2-phosophoglycollate (2PG) is also shown. A) Comparison between the liganded P168A (grey) and wild type TIM structures (purple). The dotted lines indicate the hydrogen bonding interactions between Glu167 and Ser96. Loop 6 is closed in both structures. B) Detail of the mode of binding of 2PG in wild type (green) and the P168A mutant (blue). The clash of O (Gly211) with Pro168 induced by ligand binding is indicated by an arrow. Both pictures were made with ICM (www.molsoft.com). The functional role of the conserved active site proline of triosephosphate isomerase Marco G. Casteleijn1#§, Markus Alahuhta2§, Katrin Groebel1, Ibrahim El-Sayed3, Koen Augustijns3, Annemie Lambeir4, Peter Neubauer1, Rik Wierenga2. 1) University of Oulu (Finland), Bioprocess Engineering Laboratory, Dept. of Process and Environm. Engineering and Biocenter Oulu; 2) University of Oulu (Finland), Dept. of Biochemistry and Biocenter Oulu; 3) University of Antwerp (Belgium), Laboratory of Medicinal Chemistry; 4). University of Antwerp (Belgium), Laboratory of Medicinal Biochemistry. §These authors contributed equally to this work; # corresponding author: marco.casteleijn@oulu.fi. Introduction Triose phosphate isomerase (TIM) is a glycolytic enzyme with very high substrate specificity. TIM catalyses the interconversion of D-glyceraldehyde-3-phosphate (DGAP) and dihydroxyacteone phosphate (DHAP), as seen in Figure 1. The wild type enzyme is a dimer, and each subunit has the classical TIM-barrel fold. Our studies are focusing on understanding the catalytic properties of the Wild Type (Wt-TIM) [1, 2] and applying this knowledge to our newly engineered enzymes. Enzymology of P168A vs. wild type Structural information Wild Type TIM is well studied, and many publications can be found on its catalytic and structural properties. However, some aspects are still poorly understood. Observations in a liganded structure of Leishmania TIM showed P168 (N-terminal hinge) in a planar state which is a strained[2] conformation, which could facilitate loop opening. Furthermore, proline 168 is fully conserved in all TIM species. We investigated the enzymological properties of wild type TIM and its P168A mutant regarding the loop-6/loop-7 mechanism. The results are summerized in figure 2, where the impact on conversions per second and affinity for 2PG (see also Figure 1) can be seen. Interestingly the affinity for the phosphate moiety of the substrate (see figure 1) is not affected. The results clearly show the importance of proline 168 for catalytic activity. This investigation, and of others [3, 4, 5], are of importance for protein engineering efforts on TIM, since only in the closed form the active site is competent for catalysis. This knowledge can then be applied to engineer (more) active monomeric enzyme variants (based on ml8b TIM and our own engineered A-TIM) regarding poorly binding (new) ligands. Structural investigation of liganded (with 2PG) and unliganded P168A mutant revealed two new observations. As seen in figure 3A, loop-6 and loop-7 are in the closed conformation as in wild type. However, the conformational switch from “swung out” to “swung in” of the catalytic glutamate (Glu167) has not occurred upon ligand binding in the P168A mutant. This clearly affects the mode of binding of the transition state analogue were the mode of binding is “down”; unlike in wild type (Figure 3B). A B The second new observation is that the trigger for loop-6 closure is induced by the interactions of the phosphate moiety of the ligand with residues in loop-7. This switch of loop-6 is facilitated, or even triggered, by the movement of O(Gly211) in loop-7 as it clashes with CD(Pro168). This occurs if the Pro168 side chain would not move on ligand binding (Fig. 3B). This switch does not exist in the P168A mutant. Conclusions • Proline 168 is needed for conformational strain around the catalytic glutamate. This strain is transferred to the catalytic glutamate. • Ligand binding triggers the conformational switch of the catalytic machinery needed for catalysis. References Acknowledgements This work is supported by the Academy of Finland (project 53923) an Tekes project “Biocatnuc” We have much appreciated the help from the staff at the beam-line BW7A (EMBL/DESY in Hamburg) for collecting the data of the liganded P168A structure. The skillful technical support from Ville Ratas has been invaluable in this work. • Kursula, I., and Wierenga, R.K. (2003) Crystal structure of triosephosphate isomerase complexed with 2-phospho-glycolate at 0.83Å resolution. J. Biol. Chem 278, 9544-9551. • Kursula, I., Salin, M., Sun, J., Norledge, B.V., Haapalainen, A.M., Sampson, N.S., and Wierenga, R.K. (2004) Understan-ding protein lids: structural analysis of active hinge mutants in triosephosphate isomerase. PEDS (submitted). (about importance of A178L) • Norledge, B.V., Lambeir, A.M., Abagyan, R.A., Rottmann, A., Fernandez, A. M., Filimonov, V.V., Peter, M.G., Wierenga, R.K. 2001. Modelling, mutagenesis and structural studies on the fully conserved phosphate binding loop (loop-8) of triosephosphate isomerase: towards a new substrate specificity. Proteins 42, 383-389. • Schliebs, W. Thanki, N., Eitja, R., Wierenga, R.W. (1996) Active site properties of monomeric triosephosphate isomerase (monoTIM) as deduced from mutational and structural studies. Prot. Science 5, 229-239. • Thanki, N., Zeelen, J.Ph., Mathieu, M, Jaenicke, R, Abagyan, R.A.A., Wierenga, R.K., Schliebs, W. (1997) Protein engineering with monomeric triophosphate isomerase (monoTIM): the modelling and structure verification of a seven-residue loop. Protein Engin. 10(2), 159-167.