Download
1 / 35
1.02k likes | 3.45k Views
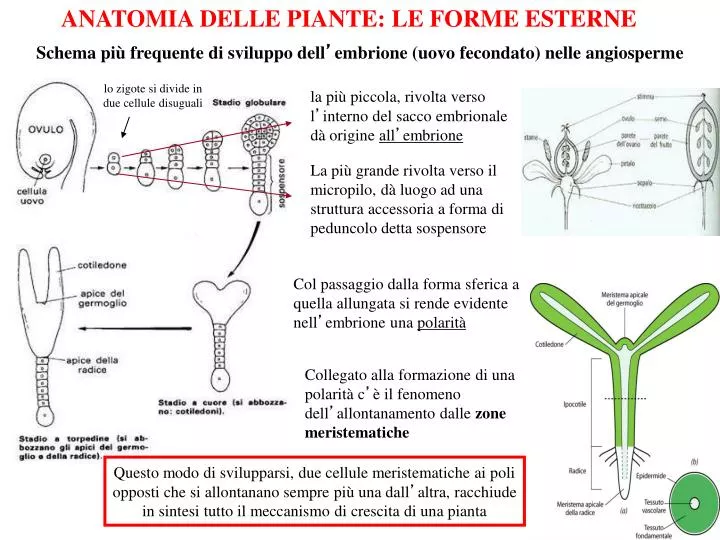
ANATOMIA DELLE PIANTE: LE FORME ESTERNE. Schema più frequente di sviluppo dell ’ embrione (uovo fecondato) nelle angiosperme. lo zigote si divide in due cellule disuguali. la più piccola, rivolta verso l ’ interno del sacco embrionale dà origine all ’ embrione.

E N D
ANATOMIA DELLE PIANTE: LE FORME ESTERNE Schema più frequente di sviluppo dell’embrione (uovo fecondato) nelle angiosperme lo zigote si divide in due cellule disuguali la più piccola, rivolta verso l’interno del sacco embrionale dà origine all’embrione La più grande rivolta verso il micropilo, dà luogo ad una struttura accessoria a forma di peduncolo detta sospensore Col passaggio dalla forma sferica a quella allungata si rende evidente nell’embrione una polarità Collegato alla formazione di una polarità c’è il fenomeno dell’allontanamento dalle zone meristematiche Questo modo di svilupparsi, due cellule meristematiche ai poli opposti che si allontanano sempre più una dall’altra, racchiude in sintesi tutto il meccanismo di crescita di una pianta
Meccanica di crescita delle piante e degli organi Modello molto semplificato di pianta sul quale evidenziare i principi fondamentali della crescita Embrione di forma sferica fatto interamente di cellule meristematiche, cioè capaci di dividersi. Ciascuna delle cellule dell’embrione si divide sempre secondo un piano parallelo all’equatore. • Questo modello simula l’accrescimento dell’asse principale (fusto + radice) • le due opposte zone meristematiche si allontanano sempre di più l’una dall’altra restando quindi separate da masse sempre più grandi di cellule adulte. • i due apici meristematici cambiano continuamente posizione rispetto al livello del terreno. • il numero totale di cellule dell’organismo aumenta continuamente ma il numero di cellule in ciascun apice non aumenta. • in qualsiasi momento dello sviluppo c’è una riserva di cellule meristematiche pronte a dividersi.
Passando dal modello ad una pianta, sia nel fusto sia nella radice si incontrano le zone di sviluppo: • Zona meristematica. • Zona di distensione e differenziamento • Zona di struttura primaria. • Zona di struttura secondaria. Meristemi in struttura secondaria: Le nuove cellule formate dal cambio cribro-vascolare si differenziano negli elementi dei due tessuti conduttori: libro e legno. In sezione trasversale libro e legno appaiono come due anelli concentrici: legno all’interno e libro all’esterno, l’anello di legno è più grosso di quello del libro. Il cambio subero-fellodermico si forma in posizione più periferica rispetto al cambio cribrovascolare. Il fellogeno genera verso l’esterno il sughero (tessuto tegumentale), mentre verso l’interno origina il felloderma (tessuto parenchimatico).
Durante la crescita del fusto le prime tre zone (meristematica, di distensione, di struttura primaria) si spostano sempre più verso l’alto (verso il basso nella radice), ma la loro estensione rimane costante. Invece la zona di struttura secondaria diventa sempre più estesa perchè il suo limite superiore si sposta verso l’alto mentre il suo limite inferiore, che coincide con la base del fusto, non cambia posizione.
Organi di una giovane pianta Dicotiledone Foglie opposte Fascio collaterale chiuso Foglia bifacciale Fascio collaterale aperto ad eustele struttura primaria Fascio radiale tetrarca struttura primaria Apparato radicale allorrizico
La radice deriva dallo sviluppo del polo radicale di embrione. Organo sotterraneo della pianta ad accrescimento geotropicamente positivo. Aspetto assile e filiforme, con accrescimento indefinito. Presente nelle Cormofite RADICE RADICE • FUNZIONI: • Fissare la pianta al substrato • Assorbire H2O, Sali minerali • Ruolo di riserva e nutritivo • Sintesi di composti organici • Trasformazione di N organico
Si suddivide in 4 zone: • meristematica (embrionale) • differenziazione • struttura primaria • struttura secondaria La radice principale si accresce verso il basso. Le radici secondarie si accrescono con ramificazioni laterali.
ZONA MERISTEMATICA: costituita da cellule embrionali primarie apicali.
ZONA MERISTEMATICA • L’apice meristematico è ben protetto da un rivestimento parenchimatico chiamato cuffia. • E’ fatto di cellule generate dal tessuto meristematico primario. • Dopo un breve periodo di vita, degenerano e si trasformano in una massa viscida chiamata mucigelche ha la funzione di diminuire l’attrito tra radice e terreno. • La cuffia non ha solo funzione di protezione, ma è anche un organo di senso che percepisce lo stimolo della gravità e fa crescere la radice verso il basso. • Le cellule meristematiche generano tutti i tessuti della radice e contemporaneamente danno origine alla cuffia. • L’apice della radice è ben strutturato: le cellule che lo formano sono disposte su file ordinate, pronte alla divisione, all’accrescimento e alla differenziazione.
ZONA DI DIFFERENZIAZIONE • Nella zona di differenziazione le cellule diventano adulte. • Nella radice questa zona è corta poiché la radice deve penetrare nel terreno duro. Serve quindi una struttura sufficientemente rigida, se questa zona che è tenera e flessibile fosse troppo lunga, la radice si piegherebbe di fronte alla resistenza offerta dal terreno e non potrebbe avanzare.
ZONA DI STRUTTURA PRIMARIA E’ la zona deputata principalmente all’assorbimento di acqua e sostanze inorganiche in essa disciolte. Il rizoderma è privo di cuticola e quindi è permeabile all’acqua ed alle soluzioni saline del terreno. Dal rizoderma si sviluppano peli unicellulari (zona pilifera) lunghi e flessibili che penetrano negli spazi capillari del terreno e assorbono acqua e sali. Secernono attivamente ioni H+ che vengono scambiati con cationi nel terreno. I peli hanno vita breve, solo pochi giorni. Le velocità di formazione e di degenerazione sono uguali per cui l’area della zona pilifera resta costante nel tempo. Segue il cilindro corticale o corteccia, il cui primo strato, che ha funzione protettiva, viene chiamato esoderma. Piùinternamente c’è un parenchima corticale per condurre in direzione radiale H2O e soluzioni. Lo strato più interno della corteccia è endoderma.Hafunzione di barriera selettiva, infatti mancando glispazi intercellulari, H2O e soluti sono costretti a passare attraverso il citoplasma delle cellule dell’endoderma. Al centro della radice c’è il cilindro centrale. Lo strato più esterno a contatto con l’endoderma, si chiama pericicloo strato rizogeno. E’ costituito da cellule in grado di riprendere l’attività meristematica e di generare radici laterali. Il cilindro centrale contiene i tessuti conduttori immersi in un parenchima fondamentale.
Rizoderma Il rizoderma è formato da cellule dell’epidermide radicale. E’ permeabile all’acqua e alle soluzioni saline del terreno. Avvolge il corpo primario della radice solo nel breve tratto retrostante la zona apicale (in genere pochi cm) formando la zona pilifera. I peli radicali sono estroflessioni delle cellule epidermiche che servono ad aumentare l’area assorbente della radice. Le dimensioni della zona pilifera si mantengono costanti nel tempo. Nella zona retrostante la zona pilifera, il rizoderma degenera e la funzione di tessuto tegumentale ad azione protettiva è assunta dall’esoderma. L’esoderma è lo strato superficiale della corteccia, le cellule (di tipo parenchimatico) suberificano progressivamente le loro pareti, formando uno strato pluricellulare simile al sughero (ma di origine primaria).
Zona pilifera La zona pilifera inizia nella zona di differenziazione e si sviluppa fino nella zona di struttura primaria della radice. Le dimensioni della zona pilifera si mantengono costanti nel tempo in ogni specie: per ogni cellula pilifera distante dall’apice ogni giorno degeneri e muoia (la vita media dei peli si misura in giorni) altrettante di nuove ne vengono differenziate dal protoderma nella zona di differenziazione. Con la sua continua crescita la radice sposta la zona assorbente in avanti nel terreno, in zone ancora non sfruttate.
Cilindro corticale Il cilindro corticale o corteccia costituisce la porzione più rilevante della struttura primaria della radice ed è formata principalmente da tessuto parenchimatico di riserva. La corteccia è delimitata nella parte esterna dall’esoderma e nella parte interna (verso cilindro centrale) dall’endoderma. L’esoderma è il tessuto tegumentale che delimita il corpo primario della radice. L’esoderma, sostituendo il rizoderma, non ha funzione assorbente, ma una volta completata la suberificazione funzionerà come tessuto tegumentale. L’endoderma è posto nella parte più interna della corteccia. E’ formato da un unico strato di cellule tegumentali (di derivazione primaria) privo di spazi intercellulari, ed ha funzione di barriera selettiva. Le pareti radiali delle cellule formanti l’endoderma sono impregnate con suberina, formando così la banda del Caspary che rende impermeabile l’accesso ai soluti.
Endodermaformato damonostrato cellulare con caratteristici ispessimenti di suberina. I punti suberificati di contatto tra le cellule sono detti punti del Caspary. Le cellule dell’endoderma esplicano funzione protettiva controllando l’assorbimento minerale: obbligano acqua e i soluti assorbiti dalla radice a passare all’interno del loro citoplasma prima di raggiungere il cilindro centrale e quindi il sistema conduttore.
Nel parenchima corticale acqua e soluti possono muoversi attraverso tre strade distinte: a) Apoplastica: spazi intercellulari, b) Simplastica: pareti cellulari, c) Transcellulare: citoplasma delle cellule. A livello delle cellule endodermiche, le bande del Caspary induco l’H2O e soluti ad attraversare la cellula (transcellulare). Avviene così il filtraggio dei soluti estranei.
Cilindro centrale Al cilindro corticale seguendo verso l’interno si sostituisce il cilindro centrale. Lo strato più esterno a contatto con l’endoderma, si chiama periciclo o strato rizogeno, ed è fatto di cellule che possono riprendere l’attività meristematica e da cui si generano le radici laterali. Il cilindro centrale contiene i tessuti conduttori, immersi in un parenchima fondamentale. Nella radice, legno e libro non sono riuniti in fasci conduttori, ma in un unico fascio vascolare a struttura raggiata -actinostelica Il fascio che contiene i tessuti conduttori è formato da ARCHE. Le arche sono formate da cordoni di tessuto di conduzione e prendono il nome di arche legnose (Xilema) e liberiane (Floema). Le arche legnose e quelle liberiane si alternano lungo la circonferenza del fascio. Il numero delle arche varia a seconda del genere e della specie (importanza sistematica).
Il cilindro centrale è composto prevalentemente da tessuto conduttore. Lo xilema e il floema formano un UNICO FASCIO a struttura raggiata con arche xilematiche e arche floematiche alternate tra di loro, che confluiscono al centro (actinostele). - Nelle arche xilematiche si distingue: protoxilema nella parte più esterna (esarco) costituito da elementi vascolari con piccolo diametro, metaxilema, nella parte più interna (endarco), di cui fanno parte grosse trachee. - Anche nelle arche floematiche il protofloema é esarco e il metafloema endarco, tuttavia le due zone sono difficilmente distinguibili.
Formazione radici laterali Le radici laterali si formano all’interno dell’organo radicale in seguito alla proliferazione di un gruppo di cellule del periciclo che si differenziano e riprendono a moltiplicarsi. Una serie di divisioni periclinali porta alla formazione di un apice vegetativo, strutturalmente e funzionalmente simile a quello della radice principale. L’apice vegetativo ha origine in prossimità di un arca legnosa (xilematica): sono così favoriti i collegamenti col tessuto conduttore. Le cellule neo-formate iniziano a distendersi. La radice così abbozzata si allunga lacerando il parenchima corticale, spingendosi verso l’esterno. Inizialmente l’endoderma segue l’accrescimento della radice laterale lungo la corteccia, poi viene lacerato quando la radice laterale esce all’esterno.
La formazione delle radici laterali ha origine dallo strato rizogeno del periciclo. Le cellule del periciclo in prossimità delle arche legnose (xilematica) riacquistano le caratteristiche meristematiche. Le cellule neo-formate iniziano a distendersi e si allungano in senso laterale. Per mezzo di enzimi litici perforano la corteccia della radice madre ed escono verso l’esterno.
ZONA DI STRUTTURA SECONDARIA E’ presente solo nelle piante che hanno una struttura secondaria (dicotiledoni). Il cambio cribro-vascolare si forma in parte dal periciclo in corrispondenza delle arche legnose, in parte da cellule parenchimatiche presenti tra le arche. L’anello cambiale appena formato avrà un aspetto sinuoso, circondando le arche legnose e lasciando fuori, verso l’esterno quelle liberiane. Questa forma sinuosa dura poco però: infatti nel primo periodo di attività il cambio produce legno secondario solo nelle zone tra le arche di legno primario; quindi il cambio viene spinto fuori ed assume una forma circolare. Successivamente il cambio lavorerà formando xilema verso l’interno e floema verso l’esterno. Il primo fellogeno può formarsi in posizione variabile, molto spesso nel periciclo. Sembra accertato che le radici mantengano una certa capacità di assorbire acqua anche nella zona di struttura secondaria dove sono rivestite dal sughero.
Crescita secondaria della radice Il corpo secondario della radice è basato sulla formazione di un cambio cribro-vascolare che prende origine da cellule parenchimatiche del periciclo. Il cambio neo-formato si dispone in modo sinusoidale tra le arche xilematiche e liberiane, lasciando all’interno il legno (xilema) e ponendo verso l’esterno il libro (floema). Successivamente il cambio da sinusoidale diverrà anulare formando xilema verso l’interno e floema verso l’esterno. Al centro rimarrà la struttura alterna delle arche xilematiche. Passaggio alla struttura secondaria nella radice 1) cilindro corticale; 2) endoderma; 3) periciclo; 4)arche xilematiche; 5) arca floematica; 6) cambio; 7) legno secondario
Radice in struttura secondaria radice Nella radice come nel fusto il tessuto legnoso è più abbondante di quello floematico. La struttura istologica dei due tessuti è uguale a quella del fusto; generalmente però il legno della radice contiene meno fibre e più vasi e le cerchie annuali sono meno marcate. Come nel fusto si sviluppano raggi parenchimatici via via che il diametro delle radice si accresce. fusto L’epidermide dopo la caduta dei peli radicali viene sostituita inizialmente dall’esoderma. Successivamente quando ha inizio l’accrescimento secondario si forma il cambio subero-fellodermico che internamente genera felloderma e all’esterno il sughero. Il complesso sughero-fellogeno-felloderma è detto periderma. Presenza di lenticelle come nel fusto. Come nel fusto si forma il ritidoma che spesso si desquama.
Radice di Dicotiledone • Dall'esterno verso l'interno si distinguono:- il rizoderma, sono riconoscibili i peli radicali con funzione di assorbimento. - la corteccia, costituita principalmente da cellule del tessuto parenchimatiche con funzione di riserva (accumulano amido) • il cilindro centrale, formato soprattutto da cellule del tessuto conduttore. • Sono riconoscibili quattro arche xilematiche (tetrarca) alternate a quattro arche floematiche. • Al centro non è presente il midollo. Nel cilindro centrale di una dicotiledone, le arche conduttrici, inizialmente, sono periferiche e costituite da protofloema e protoxilema. In seguito si forma metafloema che cresce disordinatamente e metaxilema che cresce ordinatamente verso l’interno del cilindro centrale andando a saldarsi al centro con le altre arche metaxilematiche, formando una figura a stella. Manca il parenchima midollare.
Radice di Monocotiledone In evidenza il cilindro centrale di una radice con struttura primaria "invecchiata“. L'endoderma presenta degli ispessimenti ad U molto evidenti, mentre le cellule del parenchima del cilindro centrale hanno parete ispessita e lignificata. Lo xilema e il floema, attivi nella conduzione della linfa, formano il fascio con la tipica struttura raggiata (poliarca) con un numero elevato di arche xilematiche e floematiche alternate tra loro. Al centro del cilindro è distinguibile il midollo.
RIASSUNTO: Anatomia istologica della Radice - ancoramento al suolo, - assorbimento di acqua e sali minerali, - conduzione della linfa grezza verso le parti epigee. Caratteristiche strutturali: Rizoderma: formato da tessuto tegumentale con funzioni di assorbimento e protezione Cilindro Corticale: Formato principalmente da tessuto parenchimatico. E’ molto sviluppato nella radice, ha funzioni principali di riserva. Esoderma: primo strato verso l’esterno del cilindro corticale. Sostituisce il rizoderma, ha funzioni di protezione Endoderma:ultimo strato del cilindro corticale. Ha funzione selettiva. Cilindro Centrale: formato principalmente da tessuto conduttore Periciclo: primo strato del cilindro centrale: ha funzioni rizogene. Il cilindro centrale ha UN UNICO FASCIO a struttura raggiata o alterna (Actinostele). Il numero delle arche legnose e liberiane è variabile in funzione del genere e della specie. • Cilindro corticale con parenchima di riserva • Cilindro centrale con parenchima midollare • Endoderma (ultimo strato del cilindro corticale) • Floema • Xilema • Periciclo (primo strato del cilindro centrale) Esoderma (rizoderma se zona peli radicali) Ipoderma (non sempre presente) Parenchima del cilindro corticale Endoderma
Morfologia radicale La radice è la parte del cormo che deriva dallo sviluppo del polo (apice) radicale dell’embrione. La prima radice che si forma nelle Spermatofite è detta primaria (o principale). Dalla radice principale si formano radici secondarie (o di secondo ordine), terziarie, ecc. fino ad assicurare un efficace ancoramento ed una esplorazione capillare del suolo. Nelle Dicotiledoni e Gimnosperme la radice principale rimane per la durata della vita della pianta ed ha solitamente maggiori dimensioni (apparato radicale a fittone). Quando le radici laterali assumono dimensioni analoghe alla principale si ha un apparato radicale affastellato ofascicolato. Nelle Monocotiledoni la radice primaria cessa di accrescersi e muore: l’apparato radicale è dato da radici secondarie avventizie che si originano da meristemi avventizi del fusto. In casi particolari si possono formare radici avventizie sia sul fusto che sulle foglie Nelle Dicotiledoni la radice ha maggiori dimensioni (apparato radicale ALLORRIZICO). Nelle Monocotiledonila radice principale muore e dalla base del fusto si formano radici avventizie (apparato radicale OMORRIZICO)
MODIFICAZIONE DELLA RADICE Accessorie: che originano in punti determinati del caule, vengono così suddivise: Aggrappanti: particolari radici accessorie che si sviluppano dal lato del fusto a contatto di un sostegno (Edera). Colonnari o fulcranti: particolari radici aeree, che si ingrossano e si irrobustiscono come delle colonne assumendo una funzione di sostegno (Mangrovie, ficus religiosa) Caulinari: costituiscono l’apparato radicale omorrizico delle Monocotiledoni. Avventizie: radici che originano dal fusto o dalle foglie e che si formano in seguito a fenomeni traumatici quali ferite o distacco di una porzione della pianta madre (utilizzate anche per moltiplicazione vegetativa: talea, margotta …). Tuberizzate: radici fortemente ingrossate e contenenti riserve diffuse in molte piante erbacee che sopravvivono per almeno un inverno (carota, barbabietola). Può ingrossare il parenchima corticale o quello centrale. Contrattili: nelle piante con tuberi o bulbi le radici possono raggrinzirsi e distendersi per interrire e approfondirsi nel terreno. Pneumatofori: sono radici modificate che con un geotropismo negativo, si innalzano dal terreno verticalmente con la funzione di aerazione, ricchi di parenchimi aeriferi. Tipico di alcune piante che vivono in acqua o in terreni paludosi (mangrovie). Austori: sono radici con le quali le piante parassite (cuscuta) e emiparassite (vischio) succhiano la linfa delle piante ospiti. Sono provviste di un cono di penetrazione che perfora i tessuti dell’ospite fino ad arrivare ai tessuti conduttori.
Tipici delle Fabaceae (leguminose), i batteri del genere Rhizobium sono ospitati nelle radici, dove provocano infezioni, identificabili per la presenza di tubercoli. L’infestazione radicale da parte dei batteri del genere Rhizobium, si rivela vantaggiosa, perché questi microrganismi sono azotofissatori, cioè sono in grado di ricavare azoto molecolare (N2) dall’atmosfera, e di fissarlo come azoto ammoniacale (NH4+). Altri batteri poi trasformano NH4+in azoto nitroso (NO2-) e nitrico (NO3-) assimilabile da parte delle piante ospiti. La simbiosi tra batteri radicali azotofissatori e piante superiori non è esclusiva delle Fabaceae, ma presente in altri generi (Cycas, Gunnera, Dryas, Hippophae, Alnus, etc.) con altri gruppi di Procarioti, tra cui cianobatteri e attinomiceti.
La micorriza è una simbiosi che si instaura nel suolo tra piante terrestri e funghi. Le micorrize sono diffuse in quasi tutti gli habitat terrestri, e coinvolgono la maggior parte delle piante e dei funghi superiori, sia ascomiceti che basidiomiceti. Il fungo diffonde il micelio nel terreno e assorbe molecole organiche come eterotrofo saprofita; contemporaneamente parte del micelio avvolge le radici giovani della pianta ospite con un fitto reticolo. Le ife penetrano negli spazi intercellulari del rizoderma e della corteccia suberificata, senza interessare l’apice meristematico e il cilindro centrale (micorriza ectotrofica). Nella simbiosi micorrizica le ife fungine svolgono la funzione dei peli radicali, cioè assorbono acqua e soluti dal suolo e li trasmettono all’ospite. Il fungo ha il vantaggio di beneficiare dei prodotti della fotosintesi che giungono alle radici con la linfa elaborata; in cambio, oltre a facilitare l’assorbimento minerale, fornisce fattori di crescita e vitamine. In molte orchidee terrestri le ife fungine si spingono sino all’interno delle cellule parenchimatiche (micorriza endotrofica): il rapporto è così stretto che gli stessi semi, poveri di riserve, non riescono a germinare in un terreno privo del micelio.
Importanza economica-farmaceutica delle radici Le radici essendo organi di riserva, sono usate nell’alimentazione e per trarne prodotti industriali (carota, barbabietola da zucchero, rape, radici amare…) Molte radici sono utilizzate come medicamento; la F.U.I riporta: altea, genziana, ipecacuana, liquirizia, poligala, ratania, valeriana, rabarbaro, ginseng, arnica, arpagofito
Differenze anatomiche tra fusto e radice PASSAGGIO FUSTO - RADICE • Nell’apice meristematico della radice mancano bozze fogliari e primordi dei rami • Nella radice i primi elementi del legno si differenziano nella parte più esterna del cordone procambiale, nel fusto in quella più interna • L’epidermide del fusto è differenziata come tessuto di protezione per impedire una eccessiva traspirazione; l’epidermide della radice è differenziata come tessuto assorbente. • Nel fusto i tessuti conduttori sono disposti prevalentemente in periferia, nella radice sono raccolti al centro • Nella radice il limite tra corteccia e cilindro centrale è sempre netto (endoderma), nel fusto spesso non lo è. La radice ha un parenchima corticale esteso ed un cilindro centrale piccolo; nel fusto viceversa. • Nella zona di struttura primaria del fusto la disposizione dei tessuti conduttori è diversa a livello dei nodi e degli internodi. Questa complicazione manca nella radice che non ha foglie e quindi non è articolata in nodi e internodi. • Nel fusto gli abbozzi dei rami laterali si formano a livello dell’apice meristematico, negli strati più esterni (origine esogena). Le radici laterali vengono formate a livello di struttura primaria (origine endogena) e devono farsi largo tra i tessuti della radice per emergere all’esterno






















