Download
1 / 28
280 likes | 472 Views
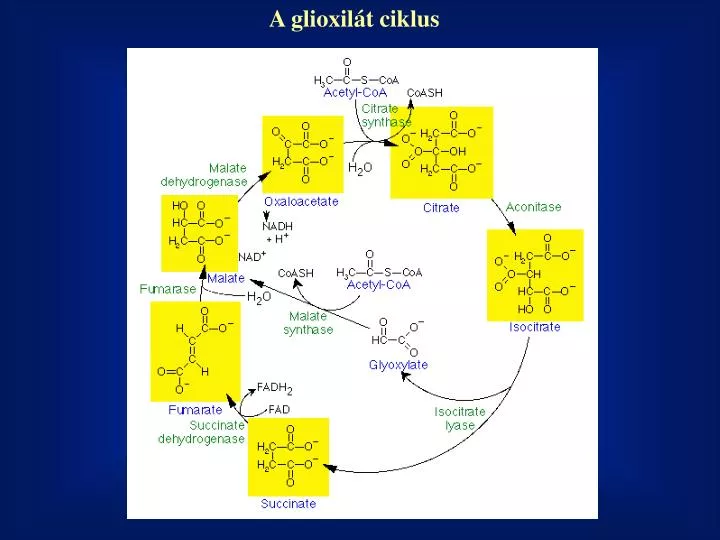
A glioxilát ciklus. Terminális oxidáció, oxidatív foszforiláció. Helye: mitokondrium belső membránja Terminális oxidáció: A kofaktorokhoz (NADH, FADH 2 ) kötött hidrogén vízzé oxidálódik. ½ O 2 + 2H + + 2e - H 2 O ½ O 2 /H 2 O E 0 = +0,82 V NADH + H + /NAD + E 0 = -0,32 V

E N D
Terminális oxidáció, oxidatív foszforiláció Helye: mitokondrium belső membránja Terminális oxidáció: A kofaktorokhoz (NADH, FADH2) kötött hidrogén vízzé oxidálódik. ½ O2 + 2H+ + 2e- H2O ½ O2 /H2O E0= +0,82 V NADH + H+ /NAD+ E0= -0,32 V DEo= 1,14 V DG0= -220 kJ/mol Oxidatív foszforiláció: ADP foszforilációja ATP-vé A terminális oxidáció és az oxidatív foszforiláció terben és időben összerendezett kapcsolt folyamatok
A mitokondriumok szerkezete Hossz: 2 mm, átmérő 0,5 mm Eredet: feltehetően aerob baktérium primitív anarob eukarióta sejt szimbiózisa Számuk fajonként és sejttípusonként változó (pl.: máj- és szívizomsejtek 800-2500 db, vvt 0) Szerkezete: kettős membrán -külső membrán: kb.: 50 % lipid, 50% fehérje, porin: átjárható a köztes anyagcsere intermedierjei számára (szelektív passzív transzport) -belső membrán: 75% fehérje, szinte valamennyi ionra nézve átjárhatatlan, a belső membrán transzporterei biztosítják a kapcsolatot a mátrix és a citoszol között. Itt található a légzési lánc elektronátvivő rendszere és az ATP-szintáz.
belső membrán mátrix FAD FADH2 O2 H2O NAD+ H+ NADH + H+ e- + + + + + + ADP Pi ATP H+ O2 + 4e- + 4H+ H2O
A mitokondriális légzési lánc alkotói, elektronszállító molekulák 4 komplex: prosztetikus csoportot tartalmazó redox folyamatokra képes fehérjék Elektrondonor: NADH + H+ , FADH2 Elektronakceptor: O2 Elektronszállító molekulák: • Citokrómok: Prosztetikus csoportként hemet tartalmaznak (Fe2+ Fe3+) abszorpciós spektrumuk alapján 3 osztályba soroljuk őket: a, b, c
2. Ubikinon: benzokinon származék,1 vagy 2 elektron felvétele/leadása. energiabomba A légzési láncban előforduló elektrontranszer szerepet ellátó prosztetikus csoportok: FeS: vas-kén komplexek. Nem hem vasat tartalmaznak: Fe2+ Fe3+ szervetlen S, vagy Cys S kapcsolódás. Réz tartalmú fehérjék: Cu+ Cu2+
redoxrendszer redoxipotenciál (V) NAD+/NADH + H+ -0,32 FAD/FADH2 -0,21 Citokrómb Fe3+/Fe2+ +0,08 Citokrómc Fe3+/Fe2+ +0,22 Citokróma Fe3+/Fe2+ +0,29 ½ O2/H2O +0,82 elektron áramlási irány
ATP Szintáz ATP ADP+Pi Az energiatermelés folyamata a mitokondriumokban porinok Inter-membrán tér H+ H+ H+ 3 H+ I e- III IV e- 3 H+ II Q Cyt c +O2 H+ H+ H+ NADH Citokróm oxidáz komplex NAD+ NADH Dehidrogenáz komplex Citokróm b-c1 komplex H2O FAD Mátrix FADH2
I. Komplex: NADH-UQ-oxidoreduktáz (NADH-dehidrogenáz) 25 polipeptidből álló fehérjekomplex. A NADH kötő hely a mátrix felé néz. Az I. komplexről az elektronok az ubikinonra kerülnek. Protonpumpa aktivitás van. II. Komplex: szukcinát-UQ-oxidoreduktáz Prosztetikus csoportja a FAD. A komplex tagja a szukcinát-dehidrogenáz (citrát ciklus). A II. komplexről az ubikinonra kerülnek az elektronok. Protonpumpa aktivitás nincs. DENAD+/NADH + H+ : UQ/UQH2 >DEFAD/FADH2 :UQ/UQH2 A FAD/FADH2 :UQ/UQH2redoxpárok közötti redoxpotenciál különbség kicsi a protonpumpa meghajtásához.
Glicerol-foszfát dehidrogenázról Zsíracil-KoA dehidrogenázról kerülhetnek még elektronok az ubikinonra
ATP Szintáz ATP ADP+Pi Az energiatermelés folyamata a mitokondriumokban porinok Inter-membrán tér H+ H+ H+ 3 H+ I e- III IV e- 3 H+ II Q Cyt c +O2 H+ H+ H+ NADH Citokróm oxidáz komplex NAD+ NADH Dehidrogenáz komplex Citokróm b-c1 komplex H2O FAD Mátrix FADH2
III. Komplex: UQH2-citokróm c-oxidoreduktáz Az elektronok az ubikinonról a citokróm c-re kerülnek. Protonpumpa aktivítás van. IV. Komplex: citokróm oxidáz Itt történik az O2 vízzé redukálása. Protonpumpa aktivítás van.
ATP Szintáz ATP ADP+Pi Az energiatermelés folyamata a mitokondriumokban porinok Inter-membrán tér H+ H+ H+ 3 H+ I e- III IV e- 3 H+ II Q Cyt c +O2 H+ H+ H+ NADH Citokróm oxidáz komplex NAD+ NADH Dehidrogenáz komplex Citokróm b-c1 komplex H2O FAD Mátrix FADH2
terminális oxidáció (elektrontranszferek sorozata) – exergonikus folyamat oxidatív foszforiláció (ADP + Pi ATP) – endergonikus folyamat Kapcsolt reakciók P/O hányados: 1 O fogyásra hány ATP szintézis jut NADH esetében: 3 FADH2 esetében: 2
ATP Szintáz ATP ADP+Pi Az energiatermelés folyamata a mitokondriumokban porinok Inter-membrán tér H+ H+ H+ 3 H+ I e- III IV e- 3 H+ II Q Cyt c +O2 H+ H+ H+ NADH Citokróm oxidáz komplex NAD+ NADH Dehidrogenáz komplex Citokróm b-c1 komplex H2O FAD Mátrix FADH2
ATP szintáz Két részből áll: F1 és Fo F1: ADP foszforilációjáért felelős rész, Fo: protoncsatorna rész szétkapcsolószerek (pl.: 2,4-dinitrofenol), a terminális oxidáció és az oxidatív foszforiláció szétkapcsolása akceptor kontroll
ATP-szintézis gátlása • F0-t oligoglicinnel eltömjük => elektrontranszfer-lánc lelassul, leáll, mert grad PH annyira megnő, hogy a deltaE már kevés a proton pumpálásához • Szétkapcsoló szerek: megszüntetik a grad pH-t, megkötik H+-t és átviszik a membrán másik oldalára • Ha nincs grad pH => nincs ATP szintézis => elektrontranszfer-lánc felgyorsul • Szétkapcsoló szerek normálisan nincsenek a szervezetben. Badibilderek eszik verseny előtt, mert jobban égetik a zsírt, ha ezzel felgyorsítják az elektrontranszfer-láncot. • Az ilyenkor termelődött energia nem tud ATP-szintézisben leadódni => hő => izzad • Szétkapcsolókat lehet hőszabályozára (pl. jegesmedvéknél téli álomkor ez termeli a hőt)
Növényeknél: • Nem lehet őket ciánnal megmérgezni, mert jobb a stressztűrő képessége, mert nem tud elmenekülni a stresszek elől. Bypassok vannak az elektrontranszfer-láncban, pl. 4 féle I. komplexe van => ha az egyik komplexet gátoljuk, a másik 3 út egyikén még mindig megy. • Alternatív oxidáz: ubikinont nem ciánszenzitíven tudja oxidálni, de ezzel is hő és nem ATP lesz. • Kb 10e db elektrontranszfer lánc / mitokondrium. Ennyi van az egyes komplexekből.
A Mitchell-féle kemiozmotikus elmélet membrán potenciál proton motoros erő: H+ ion koncentrációkülönbség






















