Download
1 / 1
10 likes | 133 Views
Action d’un herbicide, le bentazone (formulation), sur la croissance et le métabolisme d’une diatomée marine : Chaetoceros gracilis Amara A.*, Hourmant A*., Stachowski S.*, Arzul G**., Quiniou F.**, Durand G.***

E N D
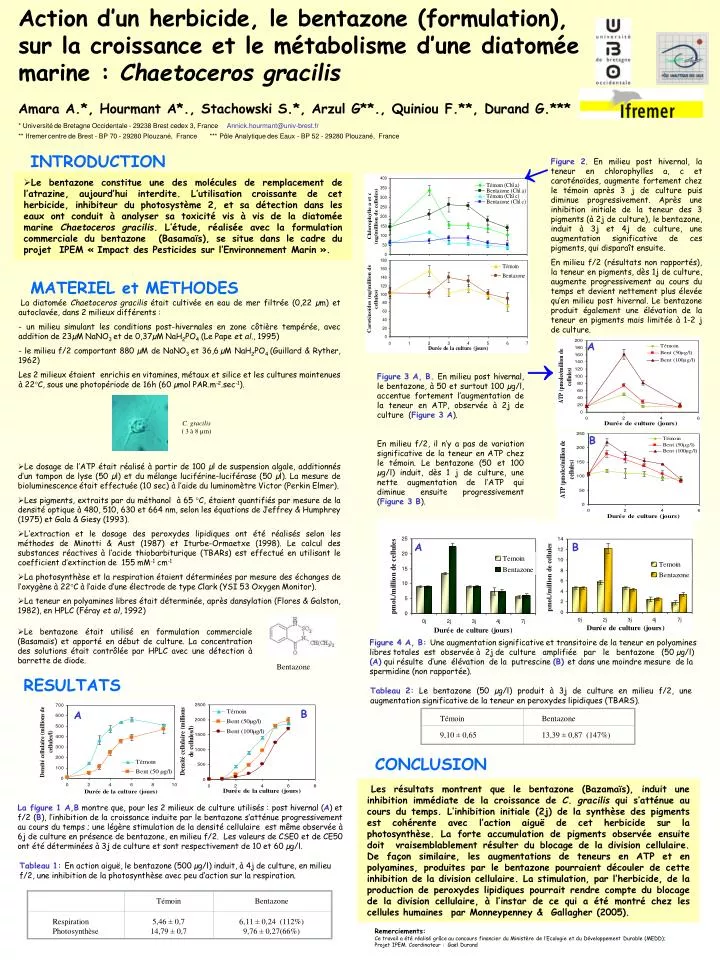
Action d’un herbicide, le bentazone (formulation), sur la croissance et le métabolisme d’une diatomée marine : Chaetoceros gracilis Amara A.*, Hourmant A*., Stachowski S.*, Arzul G**., Quiniou F.**, Durand G.*** * Université de Bretagne Occidentale - 29238 Brest cedex 3, France Annick.hourmant@univ-brest.fr ** Ifremer centre de Brest - BP 70 - 29280 Plouzané, France *** Pôle Analytique des Eaux - BP 52 - 29280 Plouzané, France Témoin Témoin Bentazone Bentazone 9,10 ± 0,65 13,39 ± 0,87 (147%) Respiration Photosynthèse 5,46 ± 0,7 14,79 ± 0,7 6,11 ± 0,24 (112%) 9,76 ± 0,27(66%) A A B B B A INTRODUCTION Figure 2. En milieu post hivernal, la teneur en chlorophylles a, c et caroténoïdes, augmente fortement chez le témoin après 3 j de culture puis diminue progressivement. Après une inhibition initiale de la teneur des 3 pigments (à 2j de culture), le bentazone, induit à 3j et 4j de culture, une augmentation significative de ces pigments, qui disparaît ensuite. En milieu f/2 (résultats non rapportés), la teneur en pigments, dès 1j de culture, augmente progressivement au cours du temps et devient nettement plus élevée qu’en milieu post hivernal. Le bentazone produit également une élévation de la teneur en pigments mais limitée à 1-2 j de culture. • Le bentazone constitue une des molécules de remplacement de l’atrazine, aujourd’hui interdite. L’utilisation croissante de cet herbicide, inhibiteur du photosystème 2, et sa détection dans les eaux ont conduit à analyser sa toxicité vis à vis de la diatomée marine Chaetoceros gracilis. L’étude, réalisée avec la formulation commerciale du bentazone (Basamaïs), se situe dans le cadre du projet IPEM « Impact des Pesticides sur l’Environnement Marin ». MATERIEL et METHODES La diatomée Chaetoceros gracilis était cultivée en eau de mer filtrée (0,22 µm) et autoclavée, dans 2 milieux différents : - un milieu simulant les conditions post-hivernales en zone côtière tempérée, avec addition de 23µM NaNO3 et de 0,37µM NaH2PO4 (Le Pape et al., 1995) - le milieu f/2 comportant 880 µM de NaNO3 et 36,6µM NaH2PO4 (Guillard & Ryther, 1962) Les 2 milieux étaient enrichis en vitamines, métaux et silice et les cultures maintenues à 22°C, sous une photopériode de 16h (60 µmol PAR.m-2.sec-1). Figure 3 A, B. En milieu post hivernal, le bentazone, à 50 et surtout 100 µg/l, accentue fortement l’augmentation de la teneur en ATP, observée à 2j de culture (Figure 3 A). En milieu f/2, il n’y a pas de variation significative de la teneur en ATP chez le témoin. Le bentazone (50 et 100 µg/l) induit, dès 1 j de culture, une nette augmentation de l’ATP qui diminue ensuite progressivement (Figure 3 B). C. gracilis ( 3 à 8 µm) • Le dosage de l’ATP était réalisé à partir de 100 µl de suspension algale, additionnés d’un tampon de lyse (50 µl) et du mélange luciférine-luciférase (50 µl). La mesure de bioluminescence était effectuée (10 sec) à l’aide du luminomètre Victor (Perkin Elmer). • Les pigments, extraits par du méthanol à 65 °C, étaient quantifiés par mesure de la densité optique à 480, 510, 630 et 664 nm, selon les équations de Jeffrey & Humphrey (1975) et Gala & Giesy (1993). • L’extraction et le dosage des peroxydes lipidiques ont été réalisés selon les méthodes de Minotti & Aust (1987) et Iturbe-Ormaetxe (1998). Le calcul des substances réactives à l’acide thiobarbiturique (TBARs) est effectué en utilisant le coefficient d’extinction de 155 mM-1 cm-1 • La photosynthèse et la respiration étaient déterminées par mesure des échanges de l’oxygène à 22°C à l’aide d’une électrode de type Clark (YSI 53 Oxygen Monitor). • La teneur en polyamines libres était déterminée, après dansylation (Flores & Galston, 1982), en HPLC (Féray et al, 1992) • Le bentazone était utilisé en formulation commerciale (Basamaïs) et apporté en début de culture. La concentration des solutions était contrôlée par HPLC avec une détection à barrette de diode. Figure 4 A, B: Une augmentation significative et transitoire de la teneur en polyamines libres totales est observée à 2j de culture amplifiée par le bentazone (50 µg/l) (A) qui résulte d’une élévation de la putrescine (B) et dans une moindre mesure de la spermidine (non rapportée). Bentazone RESULTATS Tableau 2: Le bentazone (50 µg/l) produit à 3j de culture en milieu f/2, une augmentation significative de la teneur en peroxydes lipidiques (TBARS). CONCLUSION Les résultats montrent que le bentazone (Bazamaïs), induit une inhibition immédiate de la croissance de C. gracilis qui s’atténue au cours du temps. L’inhibition initiale (2j) de la synthèse des pigments est cohérente avec l’action aiguë de cet herbicide sur la photosynthèse. La forte accumulation de pigments observée ensuite doit vraisemblablement résulter du blocage de la division cellulaire. De façon similaire, les augmentations de teneurs en ATP et en polyamines, produites par le bentazone pourraient découler de cette inhibition de la division cellulaire. La stimulation, par l’herbicide, de la production de peroxydes lipidiques pourrait rendre compte du blocage de la division cellulaire, à l’instar de ce qui a été montré chez les cellules humaines par Monneypenney & Gallagher (2005). La figure 1 A,B montre que, pour les 2 milieux de culture utilisés : post hivernal (A) et f/2 (B), l’inhibition de la croissance induite par le bentazone s’atténue progressivement au cours du temps ; une légère stimulation de la densité cellulaire est même observée à 6j de culture en présence de bentazone, en milieu f/2. Les valeurs de CSE0 et de CE50 ont été déterminées à 3j de culture et sont respectivement de 10 et 60 µg/l. Tableau 1: En actionaiguë, le bentazone (500 µg/l) induit, à 4j de culture, en milieu f/2, une inhibition de la photosynthèse avec peu d’action sur la respiration. Remerciements: Ce travail a été réalisé grâce au concours financier du Ministère de l’Ecologie et du Développement Durable (MEDD); Projet IPEM. Coordinateur : Gaël Durand






















