Download

1 / 16
361 likes | 992 Views
Mitochondria. Introduction. 1890, German scientist Altman observed mitochondria in the tissue of mouse liver and coined the term elementarorganism considering it as a living organism inside the cell.

E N D
Introduction • 1890, German scientist Altman observed mitochondria in the tissue of mouse liver and coined the term elementarorganismconsidering it as a living organism inside the cell. • Dependent on the large number of cellular instructions. There are hundred of mitochondria in the cell. • Cell's power producers converting energy into forms that are usable by the cell. • Located in the cytoplasm sites of cellular respiration, cell division and growth, as well as cell death. • In 1890 a German scientist Altman observed mitochondria in the tissue of mouse liver and coined the term elementarorganism and he thought that he found a living organism inside the cell. Today we know that mitochondria are not independent at all despite having their own genome. They relay on the large number of cellular instructions. There are hundred of mitochondria in the cell. Mitochondria are the cell's power producers. They convert energy into forms that are usable by the cell. Located in the cytoplasm, they are the sites of cellular respiration which ultimately generates fuel for the cell's activities. Mitochondria are also involved in other cell processes such as cell division and growth, as well as cell death.
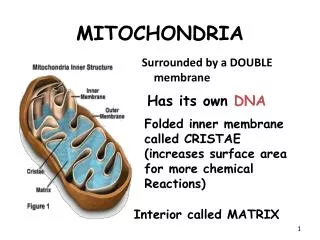
Anatomy • Shape:Discrete sausage-shaped (like meat stuffed in casings) structures • Diameter: Range from 0.5 to 10 micrometers, the second largest organelle in most animal cells • Structure: Double layered membrane organization composed of phospholipids and proteins • 5 different compartments: • Outer membrane • Inner membrane • Inner mitochondrial membrane • Cristae • Matrix
Outer membrane • Encloses entire organelle • Freely permeable to small molecule but not so significant • Has a protein-to-phospholipids ratio similar to that of the eukaryotic plasma membrane (about 1:1 by weight). • It contains large numbers of integral proteins called porins. • Porins: These porins form channels that allow molecules 5000 Daltons or less in molecular weight to freely diffuse from one side of the membrane to the other • Also has large multi subunit proteins called translocases • Translocases: ‘’carrier proteins’’ .Larger proteins can enter the mitochondrion if a signaling sequence at their N-terminus binds to a large multisubunit protein i.etranslocase of the outer membrane, which then actively moves them
Inner membrane • Permeability barrier to most solutes. • Contains 151 different polypeptides • The inner membrane is home to around 1/5 of the total protein in a mitochondrion • High protein to phospholipid ratio > (3:1) by weight. Which is about 1 protein for 15 phospholipids • Rich in phospholipids and cardiolipin • Cardiolipin:Cardiolipin contains four fatty acids rather than two and may help to make the inner membrane impermeable. • Unlike the outer membrane, the inner membrane doesn't contain porins and is highly impermeable to all molecules. Almost all ions and molecules require special membrane transporters to enter or exit the matrix. Proteins are ferried into the matrix via the translocase of the inner membrane(TIM) complex or via Oxa1. • In addition, there is a membrane potential across the inner membrane formed by the action of the enzymes of the electron transport chain.
Inner membrane Protein Functions • The inner mitochondrial membrane contains proteins with five types of functions: • Those that perform the redox reactions of oxidative phosphorylation • ATP synthase, which generates ATP in the matrix • Specific transport proteins that regulate metabolite passage into and out of the matrix • Protein import machinery. • Mitochondria fusion and fission protein • Stalk: Allows rotation of F1 complex about F0 complex • Cristae are studded with small round bodies known as F1 particles or oxysomes. These are not simple random folds but rather invaginations of the inner membrane, which can affect overall chemiosmotic function • F1F0: • F0 complex: proton translocator channel • F1 complex: proton translocator through F0 drive ATP, synthesis by F1
Matrix • The matrix is the space enclosed by the inner membrane. It contains about 2/3 of the total protein in a mitochondrion. • The matrix is important in the production of ATP with the aid of the ATP synthase contained in the inner membrane. • The matrix contains a highly-concentrated mixture of hundreds of enzymes, special mitochondrial ribosomes, tRNA, and several copies of the mitochondrial DNA genome. Of the enzymes, the major functions include oxidation of pyruvate and fatty acids, and the citric acid cycle.
Mitochondrial genome • Only location of extrachromosomal DNA with in the cell, under dual genetic control by both nuclear and mitochondrial genome. • Double stranded of about 16 kb in length. • Comprise only about 1-2% of the total DNA in the mammalian cells. • A published human mitochondrial DNA sequence revealed 16,569 base pairs encoding 37 total genes. • mtDNAencodes 13 essential proteins for OXPHOS as well as 2 rRNA and 22 tRNA which are required for the assembly of functional mitochondrial protein synthesis system. • 13 mRNA are encoded by mitochondria for 13 specific proteins. These protein interact 60 different proteins to form the enzyme complex for OXPHOS. These complexes integrate into mitochondrial membrane and are involved in the electron transport chain ATP synthesis. • The remaining 1500 mt proteins are encoded by the nucleus including all proteins needed for replication and expression of small mtDNA genome.
Mitochondrial replication machinery • The hetrotrimericPOLγtogether with mtDNAhelicase (Twinkle) and additional replisome are responsible for mtDNA replication. Antiviral nucleoside analogus are some time recognized by POLγ cause drug side effects by interfering mtDNA replication. Replication errors are responsible for age associated diseases and aging.
Carrier pathway to the inner mitochondrial membrane. The noncleavable precursors of hydrophobic metabolite carriers of the inner membrane are imported in several stages. Cytosolic chaperones guide the precursor (stage I) to the receptor Tom70 (translocase of outer membrane receptor 70) (stage II). The precursor is transported through the translocase of the outer membrane (TOM complex) in a loop formation and interacts with the Tim9-Tim10 chaperone complex (stage IIIa). Tim9-Tim10 guides the precursor through the intermembrane space (IMS) to the carrier translocase of the inner membrane (TIM22 complex with bound Tim9-10-12) (stage IIIb). The membrane potential (Δψ) promotes insertion of the precursor into the inner membrane via the TIM22 complex (stage IV) followed by assembly into the mature form of the carrier
The machinery for import and assembly (MIA): • The precursor protein with sulphide bonds come in contact with TOM 40 (translocase of outer membrane 40), after that Mia40 (mitochondrial inner membrane space assembly machinery protein) binds to the precursor via a transient disulfide bond. The sulfhydryloxidase Erv1 cooperates with Mia40 in the oxidation of the precursor protein. Erv1 reoxidizes Mia40 and transfers electrons to cytochromec .
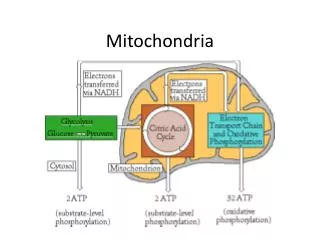
Mitochondrial energetics and Apoptosis • Under anaerobic conditions glucose is concerted into lactate and on the other way in aerobic conditions pyruvate is formed by glucose. From cytoplasm, pyuvate moves inside the mitochondria with the help of pyruvatedehydrogenase enzyme (PDH) to initiate tricarboxylic acid cycle (TCA) for the gain of energy. NADH from the TCA cycle starts ETC (electron transport chain) at Coenzyme-Q. Cytochrome C which presents in the intermembrane space plays a vital role in the ETC as well as in the programmed cell death (apoptosis). Any undesired change in the cytoplasm leads to the recruitment of Bax and Bad proteins with Bcl-2 protein. Bax, Bad and Bcl-2 protein complex punctures the outer membrane for the release of cytochrome c into the cytoplasm. In cytoplasm cyt-C make a complex with apoptosis protease activating factor 1 (Apaf 1) and caspases like C-9, C-7 and C-3 directs protein digestion and degradation of DNA results into cell death.