Download
1 / 53
530 likes | 698 Views
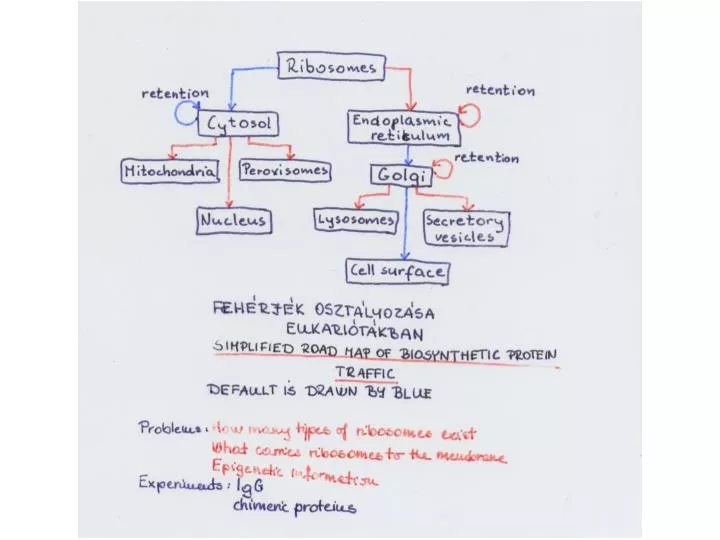
The channel through the membrane is called the translocon. Summary of transport into the ER. SRP stops translation. LOCALIZATION OF MEMBRANE-BOUND PROTEINS IN ER.

E N D
Hsp40 binds the substrate and then Hsp70. ATP hydrolysis drives conformational change. GrpE displaces the ADP; this causes the chaperones to be released. Multiple cycles of association and dissociation may occur during the folding of a substrate protein
ER proteins are glycosilated Dolichol is a lipid that consists of a long chain of isoprenoid units and is present in the membrane of the rough endoplasmic reticulum. It is part of the precursor in the synthesis of N-linked oligosaccharides. An oligosaccharide is assembled onto dolichol via a pyrophosphoryl linkage, then transferred to particular asparagine residues of a nascent polypeptide. A high mannose oligosaccharide is an N-linked oligosaccharide that contains N-acetylglucosamine linked only to mannose residues. It is covalently added to transmembrane proteins in the rough endoplasmic reticulum and is trimmed and modified in the Golgi apparatus
A high mannose oligosaccharide is an N-linked oligosaccharide that contains N-acetylglucosamine linked only to mannose residues. It is covalently added to transmembrane proteins in the rough endoplasmic reticulum and is trimmed and modified in the Golgi apparatus
A complex oligosaccharide is generated in those cases in which further residues are added in the Golgi
Lys-Asp-Glu-Leu (KDEL in single letter code).. If this sequence is deleted, or if it is extended by the addition of other amino acids, the protein is secreted from the cell instead of remaining in the lumen
Tim17-23 complex translocates proteins to the lumen The Tim22-54 complex translocates proteins that reside in the inner membrane
PEROXISOMES Consensus PTS1 tripeptides (S,A,C/K,R,H/L) interact strongly in the PTS1-binding site, without the need for additional interactions between the receptor and the PTS1-containing protein. Non-consensus PTS1 tripeptides function as targeting signals only in combination with additional interactions with the PTS1 receptor.
NUCLEAR TRANSPORT The key to understanding how directionality of nuclear transport is conferred lies in the localization of the RanGAP and RanGEF. As, RanGAP is located in the cytoplasm on the surface of the nuclear pore complex. This means that in the cytoplasm, Ran is in its GDP-bound form because RanGAP will have stimulated hydrolysis of the bound GTP. In contrast, the RanGEF is located in the nucleus. This means that in the nucleus, Ran is in its GTP-bound form. Thus, the concentration of Ran-GDP is higher in the cytoplasm and the concentration of Ran-GTP is higher in the nucleus.
Botulinum toxin: Mechanism of action at presynaptic nerve terminals2 Overall action: Blocks neurotransmitter release at peripheral cholinergic nerve terminals Neuromuscular junction Autonomic nerve terminals: Sympathetic & Parasympathetic
Synaptic vesicles are docked at the presynaptic plasma membrane (1) by SNARE complexes, associating the synaptic vesicle proteinVAMP-2 (v-SNARE) and syntaxin-1 and SNAP-25, two presynaptic membrane proteins (t-SNAREs).A fusion pore forms and opens after activation by calcium (2), allowing the release of small soluble molecules from the synaptic vesicle in the synaptic cleft. The fusion pore could be a dimer of two opposingV-ATPase membrane domains (V0 V-ATPase). The pore either expands leading to membrane fusion (3), and the full release of synaptic vesicle contents, regardless of their size. Alternatively, the pore closes after some delay (4), its transient opening being sufficient to release the synaptic vesicle neurotransmitter (NT). V-ATPase activates (5) and accumulates protons allowing neurotransmitter uptake (6).When neurotransmitter concentration is achieved, the vesicular transporter no longer dissipates the electrochemical gradient, which reaches maximal values. V-ATPase activity is turned off by dissociation of the catalytic sector V1 (7). The V0 domain is now unmasked and ready to form the V0-V0 fusion pore. This would explain how the release machinery knows that synaptic vesicles are filled with neurotransmitter and ready for release.
DCV exocytosis viewed as sequential stages of docking, priming and fusion. Kinetic components of capacitance for DCV exocytosis have been interpreted in terms of a model [14] in which vesicles transition through sequential stages. Cytoplasmic or undocked vesicles undergo docking (1). Priming of docked vesicles may occur in two stages to generate SRP (slowly releasable pool) and RRP (rapidly releasable pool) of vesicles (2 and 3). The first priming step requires moderate increases in cytoplasmic Ca2+. Activating levels of Ca2+ trigger fusion of vesicles from either the SRP (not shown) or from the RRP (4). Each step is thought to be reversible although they are depicted as single arrows in the figure. Approximate forward rate constants [14] for each step are 0.003 s1 (1), 0.03 s1 (2), 0.3 s1 (3) and 30 s1 (4). Priming reactions are thought to involve assembly of trans SNARE complexes, which may initiate by anchoring N-terminal domains of SNARE motifs (2) and their subsequent ‘‘zippering’’ to C-terminal, membrane proximal regions (3). In SV exocytosis, complexins may stabilize trans SNARE complexes at a late stage of assembly rendering them fusion-ready upon Ca2+ influx.
The interaction between SNAREs. The v-SNARE is a transmembrane protein carried by the vesicle. The t-SNARE includes two proteins; syntaxin is a transmembrane protein, and SNAP-25 is connected to the membrane by a fatty acyl linkage. (The name of SNAP-25 has an independent origin, and it has no connection with the SNAPs of the fusion particle.) Homologues to these SNAREs are found in other systems, including other animal cell types and yeast cells