Download

1 / 29
370 likes | 1.66k Views
UNIVERSIDAD DE SANTANDER UDES MEDICINA VETERINARIA INMUNOLOGIA Y VIROLOGIA VETERINARIA. EL SISTEMA DEL COMPLEMENTO. EL SISTEMA DEL COMPLEMENTO.

E N D
UNIVERSIDAD DE SANTANDER UDES MEDICINA VETERINARIA INMUNOLOGIA Y VIROLOGIA VETERINARIA EL SISTEMA DEL COMPLEMENTO
EL SISTEMA DEL COMPLEMENTO • La inmunidad innata cumple un papel de erradicación de los agentes patógenos gracias a procesos humorales y celulares. En el caso de la inmunidad humoral no específica, se puede mencionar el sistema del complemento.
EL SISTEMA DEL COMPLEMENTO • Es un sistema de defensa y limpieza , constituido por una serie de proteínas solubles, transportadas por la sangre, que une de manera covalente proteínas específicas a la superficie microbiana. • Proteínas plasmáticas (opsonizar e inducir respuesta inflamatoria.
EL SISTEMA DEL COMPLEMENTO • Puede ser activado por Ag luego de reaccionar con Acs formando complejo Ag-Acs, al hacerlo sus proteínas se solidifican y adhieren a la superficie de las células, gérmenes o moléculas que deben ser destruidas liberando simultáneamente moléculas pequeñas que incrementan la fagocitosis y amplían los mecanismos de inflamación.
EL SISTEMA DEL COMPLEMENTO • Está formado por unas 30 glucoproteinas y fragmentos que se encuentran en el suero y otros líquidos orgánicos de forma inactiva. • Su nomenclatura C1, C2, C3, C4, C5, C6, C7, C8, C9..... • Factores B, D, H e I, properdina (P). • Lectina enlazante de manosa (MBL), asociadas a serinproteasa MASP1 – MASP2. • C1 inhibidor (C1-INH, serpin), C4-enlazante de proteínas (C4-BP), factor de declinación acelerada (DAF), receptor 1 del complemento(CR1), proteína-S (vitronectina).
PRODUCCION DEL SISTEMA COMPLEMENTO. • Hígado (hepatocito):FactoresC3, C6 y C9 y el inhibidor del C1, así como el properdín y el factor B de la vía alterna. • Bazo:C6 y el C8. • Macrófagos: C4 y el C2. • Células epiteliales del intestino: C1 • Tejido adiposo: factor D • Fibroblastos:C2, C3, C5 y C9. • Riñón: C3 y C4. • Neumocitos: C3,C9.
FUNCIONES DEL SISTEMA COMPLEMENTO. • Actúa como opsonina para los fagocitos. • Induce liberación de mediadores de inflamación. • Destruye bacterias y células por acción lítica sobre las membranas. • Atrae PMN al lugar de la agresión. • Procesa e inactiva complejos Ag–Ac. • Activa LB. • Activa los fagocitos y NK • Coadyuvante en la limpieza de células apoptósicas.
FUNCIONES DEL SISTEMA COMPLEMENTO. • El objetivo básico del sistema es lograr la deposición de fragmentos del complemento sobre los blancos patológicos para la opsonización, lisis y liberación de péptidos que promuevan la respuesta inflamatoria, éste participa en la regeneración y reparación fisiológica de los tejidos.
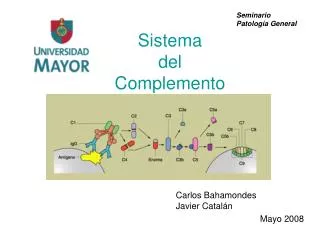
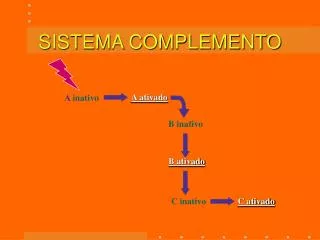
ACTIVACION DEL COMPLEMENTO • Es un proceso repetitivo, en el cual intervienen enzimas tipo proteasas de serina, existen diferentes vías que convergen en la activación de su componente central: C3. • Vía clásica. • Vía alterna. • Vía lectinas.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • Se inicia tras la unión Ag‑Ac y siempre que el anticuerpo que participe en ello sea del tipo IgM o IgG. • La interacción entre estos suscita cambios en la conformación molecular del fragmento Fc de la inmunoglobulina. Estos cambios generan un sitio de adhesión para C1 expuestos en la sección CH2 de la partícula del anticuerpo.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • La molécula de C1 está formada por subunidades C1q (que posee seis brazos helicoidales y un tallo), 2 subunidades C1r y 2 subunidades C1s. • Se desarrolla la interacción complejo anticuerpo-antígeno + cabezas globulares de la estructura helicoidal de C1q. Se requiere, por lo menos, la participación de dos sitios de unión a C1q. • La IgM por su parte, expone más sitios de adhesión. Esto explica el porqué la Ig M es más propensa a activar el sistema.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • La interacción entre la región Fc y C1q activa a C1r adquiriendo propiedades de serinproteasa, C1r . • C1r por su parte, clivará a C1s generando su forma activa: C1s. • C1s tiene dos sustratos, C2 y C4. Será primero C4 quien interactúe con él produciendo C4a yC4b . C4b se fijará a la membrana de la célula blanco, C4a difunde hacia el plasma.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • C2 por su parte, encuentra un sitio de unión en C4b, como todo este complejo se encuentra cerca de C1s , C2 es clivado por ésta generando C2a y C2b. En este caso, el fragmento más pequeño es C2b (que difunde hacia el plasma) quedando unido a C4b el fragmento C2a. • Queda formado el complejo C4b2a (C4b + C2a). A éste se le denomina "Convertasa de C3", porque justamente C3 es su sustrato a quien activa generando C3a y C3b.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • La convertasa C4b2a es capaz de generar decenas de moléculas de C3b, por ello se le suele llamar a este paso "amplificador". • C3b se unirá a C4b2a generando C4b2a3b, complejo que recibe el nombre de "Convertasa de C5". La subunidad C3b de este complejo se unirá a C5 para que el resto lo hidrolize generando C5a y C5b. C5b es un componente clave para la formación del "sistema de ataque" a la membrana celular.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • Participan aquí C5b, C6, C7, C8 y C9 con la finalidad de formar una estructura molecular conocida como complejo de ataque a la membrana. Éste se insertará en la misma a fin de permitir el ingreso masivo de iones, partículas y agua provocando la pérdida de la estabilidad celular y su muerte.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • C5b se une a la membrana celular, siempre en la región hidrófila externa de esta. • C5b se une a C6 formando C5b6. • C5b6 interactúa con C7 formando un complejo que comienza a exponer regiones hidrófobas capaces de penetrar a la sección interna de la bicapa lipídica, el complejo recibe el nombre de C5b67. • C8 se une a C5b67 formando un pequeño poro de 10 amstrong capaz de destruir glóbulos rojos pero no células con núcleo.
VIA CLASICA DE ACTIVACION DEL COMPLEMENTO • Finalmente, varias unidades C9 se unen al complejo C5b678 generando un poro que mide entre 70 y 100 Amstrong. • Por él puede ingresar a la célula una gran cantidad de iones, moléculas y agua a la vez que se pierden electrolitos importantes desembocando en la muerte de la misma.
VIA ALTERNA DE ACTIVACION DEL COMPLEMENTO • La vía alternativa fue originalmente descrita como el sistema de la Properdina. • Glicoproteína existente en el plasma sanguíneo, en donde interviene en la activación del complemento. Posee una actividad bactericida y desempeña un papel importante en la inmunidad natural. • Proteínas C3, factor B, factor D y Properdina. La vía alternativa no necesita anticuerpos para activarse. • Un requerimiento inicial para la activación es la presencia de C3b, el cual es generado en forma continua en cantidades pequeñas en la circulación.
VIA ALTERNA DE ACTIVACION DEL COMPLEMENTO • Esta activación se produce muy probablemente después de la ruptura inducida por agua del enlace tioester en C3, el cual reacciona con los factores B y D para generar un encima de fase liquida capaz de fragmentar a C3 en C3a y C3b. • Involucra un paso llamado de amplificación, donde el complejo 'Convertasa de C3' genera C3a y C3b en cantidad. Este C3b interactuará opsonizando células blanco o agentes patógenos.
VIA ALTERNA DE ACTIVACION DEL COMPLEMENTO • Por lo tanto la activación puede ser mediante dos mecanismos: • Activación secundaria a la actividad de la vía clásica, que genera C3a y C3b. • Por hidrólisis espontánea de C3 que forma C3a y C3b. • El C3b , se unirá en la membrana a antígenos extraños. Una vez afianzado a éstos puede servir como sitio de unión del Factor soluble B.
VIA ALTERNA DE ACTIVACION DEL COMPLEMENTO • El factor B unido a C3b genera el complejo C3bB que se encuentra estabilizado por magnesio y que sufre un clivaje por parte de otro factor llamado Factor D. Produce a partir del factor B los productos Ba y Bb, el primero difunde hacia el plasma y el segundo queda anclado en el complejo formando C3bBb. • El complejo C3bBb funciona como convertasa de C3 amplificando la reacción tal como en la vía clásica.
VIA ALTERNA DE ACTIVACION DEL COMPLEMENTO • Pero para que el complejo C3bBb funcione requiere de la unión a una proteína sérica llamada Properdina. • El C3b generado por el complejo C3bBb se une a este mismo constituyendo una molécula más grande llamada C3bBb3b, complejo análogo al C4b2a3b de la vía clásica también llamada 'Convertasa de C5‘.
VIA ALTERNA DE ACTIVACION DEL COMPLEMENTO • C3bBb3b toma por sustrato a C5 generando C5a y C5b. C5a difunde hacia el plasma y C5b se fija a la membrana de la célula blanco para continuar con el desenlace común a todas las vías con la formación del complejo de ataque a la membrana. (C6 - C7 - C8 - C9).
VIA LECITINA DE ACTIVACION DEL COMPLEMENTO • Dentro de la familia de las colectinas, se encuentra la proteína MBL la cual es capaz de unirse a un amplio espectro de hidratos de carbono. • Estos hidratos de carbono se encuentran presentes con mucha frecuencia en las superficies celulares de varios microorganismos. • La unión de MBL a estos componentes desencadena la activación de proteasas asociadas a ella llamadas MASP-1 y MASP-2.
VIA LECITINA DE ACTIVACION DEL COMPLEMENTO • Todo este complejo es similar al componente C1 que actúa en la vía clásica. (con sus subunidades C1q, C1r, C1s). • MASP-2 cumple una función análoga a C1s, toma como sustrato a C2 y C4 generando C2a, C2b, C4a y C4b.
VIA LECITINA DE ACTIVACION DEL COMPLEMENTO • Formación del complejo "convertasa de C3": C4b2a . • Amplificación del proceso, generación de más C3b y C3a. • Unión de C3b al complejo C4b2a: formación de convertasa de C5 (C4b2a3b). • Clivaje de C5 en C5a - C5b. • Unión de C5b a C6, C7, C8. Formación de un poro pequeño. • Ensamblaje de C9 y formación del poro mayor con la consecuente muerte celular.