Download
1 / 58
610 likes | 1.22k Views
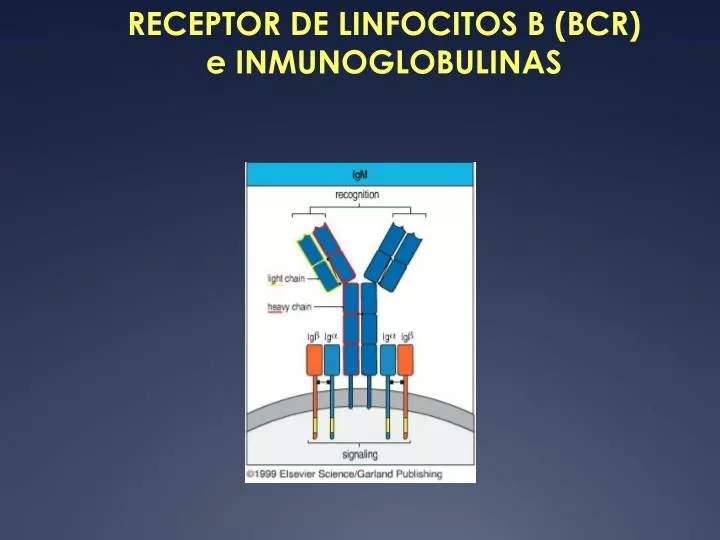
RECEPTOR DE LINFOCITOS B (BCR) e INMUNOGLOBULINAS. TEORIA DE LA SELECCIÓN CLONAL. El receptor de linfocitos B (BCR), es un complejo glicoproteico transmembrana, que incluye dos subunidades estructural y funcionalmente distintas (no covalentemente unidas entre sí).

E N D
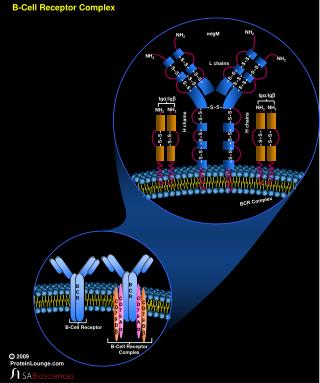
El receptor de linfocitos B (BCR), es un complejo glicoproteico transmembrana, que incluye dos subunidades estructural y funcionalmente distintas (no covalentemente unidas entre sí). • La primera subunidad es una inmunoglobulina (Ig) de membrana, que actúa como receptor clonotípico, responsable del reconocimiento específico del antígeno (Ag). • La segunda subunidad corresponde al complejo accesorio Ig/Ig, responsable del transporte y expresión de BCR en la membrana y de la transducción de señales de activación, cuando el receptor se une al epitopo antigénico.
En el repertorio linfocitario B (1014), linfocitos que reconocen distintos epitopos antigénicos tienen un receptor clonotípico propio, pero conservan el comlejo IgA/IgB, común a todos los linfocitos B. • Luego del reconocimiento antigénico, los linfocitos deben activarse y proliferar, para entrar finalmente en una etapa de diferenciación que los convertirá en linfocitos B de memoria o en células plasmáticas. • Las células plasmáticas sintetizan y secretan anticuerpos (Ac) que corresponden a la forma soluble de la Ig que tiene en su membrana los linfocitos B de los cuales derivan estas células plasmáticas.
Todas las Igs., tanto de membrana como de secreción, tienen una estructura básica constituída por 2 cadenas pesadas (H) idénticas entre sí y 2 cadenas livianas (L), también idénticas entre sí. • Cadenas H y L se asocian formando una estructura simétrica, compuesta por 2 heterodímeros H/L idénticos y covalentemente unidos entre sí por puentes disulfuro ubicados por detrás de una región “hinge” o bisagra, de gran movilidad. • En la región aminoterminal de cada heterodímero existe un paratopo o sitio de combinación para el Ag, formado por el dominio aminoterminal de cadenas pesadas y livianas. • La Ig de membrana es una molécula bivalente con 2 sitios de combinación para el Ag y monofuncional: sólo tiene función de reconocimiento antigénico. La región carboxiterminal de las cadenas H contiene una región hidrofóbica de anclaje a la membrana y un dominio citoplasmático.
En las cadenas livianas (de 25 kDa) se distingue una región o dominio variable (VL) aminoterminal de 110 aminoácidos y un dominio carboxiterminal (CL) de 110 aminoácidos. • Así, cadenas L obtenidas de Ig de membrana y anticuerpos que reconocen distintos epitopos, tendrán distinta región VL pero idéntica región CL • Sin embargo, diferencias en el dominio CL de cadenas livianas permiten distinguir dos clases, tipos o isotipos de cadenas L, denominados cadenas kappa ( ) y lambda ()
Las cadenas pesadas (de 50kDa) contienen un dominio variable (VH) aminoterminal de 110 aminoácidos y 3 o 4 dominios constantes (CH1, CH2, CH3 y CH4) carboxiterminales de 110 aminoácidos cada uno. • Variaciones en la región constante de las cadenas pesadas, permiten distinguir 5 clases, tipos o isotipos de cadenas pesadas, denominadas y (que contienen 4 dominios CH) y , , (que contienen 3 dominios CH). • La clase de cadena pesada de un anticuerpo, determina la clase, tipo o isotipo del anticuerpo. Así, - anticuerpos IgM contienen cadena pesada . - anticuerpos IgD contienen cadena pesada . - anticuerpos IgG contienen cadena pesada . - anticuerpos IgA contienen cadena pasada . - anticuerpos IgE contienen cadena pesada .
La región constante de la cadena pesada es responsable de la función efectora de un anticuerpo. Por lo tanto, las diferencias en la región constante determinan diferencias en la función efectora de los anticuerpos IgM, IgG, IgA, IgE e IgD (opsonización, activación del sistema del complemento, citotoxicidad dependiente de anticuerpos (ADCC), presencia en las secreciones, movilización transplacentaria). • Linfocitos B vírgenes, contienen en su membrana IgM e IgD que tienen especificidad por el mismo epitopo antigénico. • Linfocitos B de memoria, expresan sólo IgM, IgG, IgA o IgE. Luego del reconocimiento antigénico, estos linfocitos B de memoria darán origen a células plasmáticas que secretan IgM, IgG, IgA o IgE, respectivamente.
* En los linfocitos B, la Ig de membrana es siempre un monómero formado por 2 cadenas livianas y 2 cadenas pesadas. * Entre los anticuerpos en cambio, es posible encontrar Ig poliméricas (poli-Ig) que forman dímeros, trímeros, pentámeros o hexameros. * Así, un anticuerpo de clase IgM es un pentámero o hexámero mientras IgA puede encontrarse como monómero, dímero o trímero. * La polimerización de esta poli-Ig ocurre por asociación con una proteína J (joining) de 15 kDa, que permanece covalentemente unida al extremo carboxilo de las cadenas pesadas.
La región variable de cada cadena liviana o pesada, contiene 3 regiones hipervariables (HVR) que conforman el paratopo de una inmunoglobulina. • Estas regiones hipervariables, denominadas Complementary Determinig Regions (CDRs) determinan la especificidad por el antígeno y, en dirección amino-carboxilo se designan CDR1 (ubi cada entre los aminoácidos 25 y 35), CDR2 (entre los aac, 50 y 60) y CDR3 (entre los aac. 90 y 108), respectivamente.
El receptor de linfocitos T (TcR), es un complejo glicoproteico transmembrana, que incluye dos subunidades estructural y funcionalmente distintas ( y no covalentemente unidas entre sí). • La primera subunidad es el receptor clonotípico, TcR o TcR, responsable del reconocimiento específico del antígeno (Ag). • La segunda subunidad corresponde al complejo accesorio CD3, responsable del transporte y expresión de TcR en la membrana y de la transducción de señales de activación, cuando el receptor clonotípico se ha unido específicamente al antigéno. • El receptor clonotípico es un heterodímero o , que reconoce fundamentalmente fragmentos peptídicos asociados a una molécula de presentación antigénica (moléculas MHC de clase I o MHC de clase II) o antígenos glicolipídicos asociados a moléculas CD1.
Mientras el repertorio linfocitario B puede reconocer tanto antígenos solubles como de membrana, de naturaleza proteica, hidrocarbonada, lipídica, nucleotídica, etc., el repertorio linfocitario T reconoce fundamentalmente fragmentos peptídicos asociados a molécula MHC de clase I o MHC de clase II. • En el repertorio linfocitario B (de 1014 linfocito B distintos), se distinguen dos subpoblaciones celulares distintas (denominadas B1 y B2) que presentan características estructurales y funcionales distintas y que se generan a edades distintas durante la ontogenia linfocitaria.
En el repertorio linfocitario T (1018 linfocitos T), se distinguen también dos subpoblaciones linfocitarias ( denominadas T y T), que presentan receptores TcR estructural y funcionalmente distintos. • Entre los linfocitos T se distinguen además, 3 subpoblaciones celulares distintas denominadas TCD4+, TCD8+ y NKT. Existen además linfocitos TCD4+/TCD8+ (DP =doble positi vos) y linfocitos DN (doble negativos) • Los linfocitos NKT corresponden a linfocitos T que presentan marcadores propios de células NK (molécula NK1.1 o CD161)
El 90 a 95% de los linfocitos T corresponden a linfocitos T, que reconocen fundamentalmente fragmentos peptídicos unidos a una molécula de presentación MHC, en la membrana de una célula presentadora de antígeno. • El 5 a 10% restante, corresponden a linfocitos T entre los cuales se pueden distinguir linfocitos que reconocen fagmentos peptídicos unidos a moléculas MHC, linfocitos T que reconocen glicolípidos unidos a moléculas CD1 y finalmente, linfocito T que como los linfocitos B, pueden reconocer directamente diversas moléculas (particularmente compuestos fosforilados).
Los linfocitos T se encuentran fundamentalmente en piel y mucosas (linfocitos intraepiteliales) y se generan antes (TcR1)que los linfocitos T (TcR2), durante la ontogenia linfocitaria en el timo. • El receptor clonotípico T, es un heterodímero T o T formado por dos cadenas glicoproteicas de 40 a 60 kDa, covalentemente unidas entre sí por puentes disulfuro. • En cada cadena del receptor clonotípico T o T, se distin- gue: un dominio variable (V) aminoterminal de 112 a 119 aminoácidos, un dominio constante C, de 139 a 178 amino- ácidos, un dominio hidrofóbico transmembrana de 25 amino- ácidos y una pequeña cola citoplasmática de 12 aminoácidos.
El inicio de la respuesta inmune requiere moléculas accesorias e invariantes: A. de reconocimiento B. de señalización C. de adhesión
GENETICA MOLECULAR DE INMUNOGLOBULINAS
En los linfocitos B, La expresión de una molécula de Ig está codificada por 3 familias génicas distintas: - una familia génica que codifica la cadena pesada - dos familias génicas distintas codifican la cadena liviana (una codifica la cadena y otra codifica la cadena ). • En humanos la familia génica para la cadena pesada se encuentra en el cromosoma 14 y los genes para cadenas livianas kappa y lambda, se encuentran en los cromosoma 2 y 22 respec tivamente. • En el ratón, la familia génica para la cadena pesada se encuentra en el cromosoma 12 y los genes para cadenas livianas kappa y lambda, se encuentran en los cromosoma 6 y 16 respec tivamente.