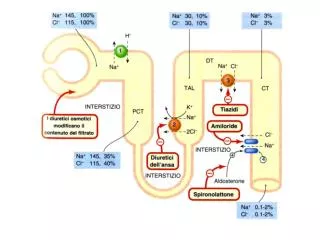
Download

1 / 15
150 likes | 414 Views
RECEPTOR DE ENZIMAS TRANSMEMBRANA, PEQUEÑAS PROTEÍNAS G, Y CASCADAS DE MAPK. Alejandro Brenes Loaiza A91047 Sección III Cap. 25. LOS RECEPTORES DE ENZIMAS TRANSMEMBRANA SON TIPOS ESPECIALES DE PROTEÍNAS.

E N D
RECEPTOR DE ENZIMAS TRANSMEMBRANA, PEQUEÑAS PROTEÍNAS G,Y CASCADAS DE MAPK Alejandro Brenes Loaiza A91047 Sección III Cap. 25
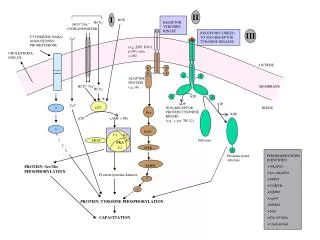
LOS RECEPTORES DE ENZIMAS TRANSMEMBRANA SONTIPOS ESPECIALES DE PROTEÍNAS Las enzimas receptoras transmembrana combinan las funciones de percepción de la señal y la actividad enzimática en el mismo polipéptido. El dominio extracelular, que recibe la señal, está conectado al dominio de la enzima intracelular por un paso único de dominio transmembrana. Las enzimas receptoras transmembrana en sistemas animales son de muchos tipos (algunos son incluso fosfatasas).
En su mayoría son quinasas de receptor de tirosina (TRKs), es decir, el aminoácido que es fosforiladopor ATP en la percepción de la señal es la tirosina. No está claro cómo un cambio en la conformación de suficiente magnitud es provocado con un solo paso de dominio transmembrana, pero la mayoría de estos receptores dimeriza al enlace ligando y una vez dimerizado, los dominios de la kinasa son capaces de fosforilarsemutuamente, amplificando así la señal intracelularmente.
En plantas, todas las quinasas de receptor transmembrana identificadas hasta la fecha con una excepción muy importante, son quinasas serina/treonina, y aunque se sospecha que la dimerización no está firmemente establecida, estos receptores de quinasas son modificaciones de los sistemas de señalización bacterianas de dos componentes que se combinan, las funciones de regulador receptor/transmisor y respuesta en un único polipéptido.
La Histidina es un receptor de quinasas, una vez más se pensó en ser restringido a procariotas, ahora son conocidos cada vez más en eucariotas, incluyendo plantas.
Receptor- como quinasas (RLKs) en plantas El número de receptor de quinasas de serina/treonina en plantas ha aumentado rápidamente desde su primera identificación en maíz en 1990. El genoma secuenciado de Arabidopsis tiene más de 300 secuencias de codificación de estas proteínas y más secuencias en otras plantas se identifican cada año.
Estos receptores de quinasas representan un tipo predominante de receptor en plantas. Sin embargo, las funciones precisas de muchos de ellos todavía no se conocen. Por lo tanto, por el momento, es apropiado referirse a ellos como receptor- como quinasas o RLKs
Los RLKs se dividen en familias en la base de sus dominios extracelulares Basada en la estructura prevista de sus dominios extracelulares, esta superfamilia de quinasas se dividen en al menos 20 familias y muchas más subfamilias. Dos motivos comunes son (I) el dominio S y (II) la repetición rica en leucina (LRR). El dominio de S se refiere a un motivo descrito por primera vez en determinadas glicoproteínas que segregan con la autoincompatibilidad (SI).
El motivo se caracteriza por una matriz de 10 residuos de cisteinaconservados adyacentes al dominio transmembrana y algunos otros residuos conservados considerados importantes en el correcto plegamiento de dominio extracelular. El motivo LRR se caracteriza por varias repeticiones en tándem de aminoácidos ricos en leucina con una secuencia de núcleo conservada Leu-x-x-Leu-x-Leu-x-x-Asnx-Leu. La repetición puede ser interrumpida por lagunas o inserciones dentro o entre repeticiones. El motivo LRR ocurre en muchas otras proteínas además de RLKstransmembranas y se piensa que median las interacciones proteína-proteína.
FIGURA 25-13Dibujos esquemáticos del receptor como quinasas (RLKs) y proteínas relacionadas con RLK. De izquierda a derecha: una glicoproteína de S-locus (SLG) con un dominio S; un RLK S-locus (SRKl); un RLK con un motivo de repetición ricos en leucina (LRR); Cf-9, una proteína que confiere resistencia a la enfermedad a un molde de hoja de tomate; PTO, una Ser/Thr quinasa que confiere la resistencia en el tomate a un patógeno bacteriano; y Pro25 con un dominio como factor de crecimiento epidérmico. Adaptado de Braun y Walker (1996) con permiso de ElsevierScience.
Las RLKsestán implicadas en Diversas funciones Estudios utilizando cruces genéticos y/o mutantes y patrones de expresión de mRNAs sugieren que RLKs tiene diversas ubicaciones y funciones celulares. Además de la respuesta de autoincompatibilidad, ellos están implicados en diversos aspectos del desarrollo de la planta y morfogénesis, incluyendo la regulación de la rama y tamaño del meristemo floral, diferenciación de células epidérmicas de hojas, abscisión, desarrollo de polen y competencia de células somáticas para formar embriones.
Por ejemplo, CLAVATAl (CLVl) es un RLK con un motivo LRR en el dominio extracelular y, desde el análisis mutacional, es conocido para regular el tamaño vegetativo y meristema floral de las ramas. Otros RLKs están involucrados en la defensa de la planta, incluyendo la resistencia a enfermedades en el arroz y el trigo, y en la regulación de respuestas de las plantas contra la tolerancia al estrés frente a la desecación, la salinidad y temperatura fría.
Las Proteínas RLK-relacionadas También son conocidas las proteínas relacionadas con RLKs. Estas son proteínas truncadas que carecen de uno o dos dominios típicos de RLKs Los ejemplos incluyen Slocus glicoproteína (SLG), que tiene un dominio extracelular similar a la de Brassica SRK, pero carece de la transmembrana y los dominios de la quinasa El PTO, una Ser/Thr quinasa que confiere la resistencia en el tomate a un patógeno bacteriano, pero carece de los dominios transmembranales y extracelulares.
La función de estas proteínas también es poco clara en muchos casos, aunque es posible que ellos formen heterodímeros con sus socios afines y función en la modulación de la señal. Un ejemplo clásico de comunicación de planta patógeno es proporcionado por PTO en tomate. PTO es una quinasa truncada serina/treonina que se produce en el citoplasma. No se sabe si dimeriza con el dominio intracelular de un socio RLK de larga duración, pero reconoce y se une al factor de antivirulencia secretada por Pseudomonas sp. que infecta y causa la enfermedad de las manchas foliares. Ese reconocimiento inicia una serie de respuestas de defensa por la planta huésped.