Download
1 / 42
420 likes | 563 Views
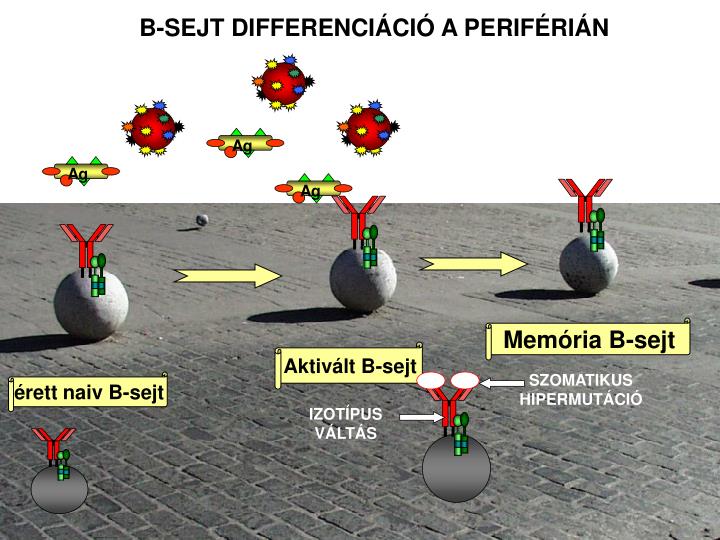
a. a. Ag. Ag. Ag. B-SEJT DIFFERENCIÁCIÓ A PERIFÉRIÁN. Memória B -sejt. Aktivált B -sejt. SZOMATIKUS HIPERMUTÁCIÓ. érett naiv B -sejt. IZOTÍPUS VÁLTÁS. Saját felismerés Klonális deléció. Saját struktúra. PERIFÉRIÁS NYIROKSZERVEK. Hozzáférhető B-sejt készlet. Antigén függő

E N D
a a Ag Ag Ag B-SEJT DIFFERENCIÁCIÓ A PERIFÉRIÁN Memória B-sejt Aktivált B-sejt SZOMATIKUS HIPERMUTÁCIÓ érett naiv B-sejt IZOTÍPUS VÁLTÁS
Saját felismerés Klonális deléció Saját struktúra PERIFÉRIÁS NYIROKSZERVEK Hozzáférhető B-sejt készlet Antigén függő Klonális osztódás Antigén - idegen Memória sejt készlet Effektor sejt készlet Potenciális B-sejt készlet CSONTVELŐ
Az immunoglobulinok molekuláris genetikája • Hogyan tud az ugyanolyan specificitású ellenanyag szekretált és membrán kötött formában megjelenni? • MEMBRÁN ÉS SZEKRETÁLT IMMUNOGLOBULIN • Hogyan képes egy adott specificitású ellenanyag egymást követően változtatni a C régió kifejeződésén? • IZOTÍPUS VÁLTÁS • Hogyan képesek az ellenanyagok növeli az antigénhez történő affinitásukat az immunválasz előrehaladtával? • SZOMATIKUS HIPERMUTÁCIÓ
Cm Primer RNS átirat AAAAA Szekretoros szakaszt kódoló szekvencia A H lánc egyes doménjeit külön exonok kódolják pA szekretált pA membrán Cm1 Cm2 Cm3 Cm4 Membrán szakaszt kódoló szekvencia A KONSTANS RÉGIÓT EXON SZAKASZOK KÓDOLJÁK
Cm1 Cm2 Cm3 Cm4 DNS Transcription pAm Cm1 Cm2 Cm3 Cm4 1° átirat AAAAA Hasítás, poly-adeniláció pAm helyen és RNShasítás Cm1 Cm2 Cm3 Cm4 AAAAA Fehérje A membrán kódoló szekvencia a transzmembrán régió révén biztosítja a sejtfelszíni expressziót Fc Membrán IgM konstant régió mRNS
h Cm1 Cm2 Cm3 Cm4 DNS Átírás pAs Cm1 h Cm2 Cm3 Cm4 1° átirat AAAAA Cleavage polyadenylation at pAs and RNA splicing Cm1 h Cm2 Cm3 Cm4 AAAAA mRNA Protein A szekréciós szakaszt kódoló szekvencia az oldott fehérje C-terminális szakaszának szintéziséért felelős Fc Szecernált IgM konstans régió
Az emberi nehéz lánc gének C régióinak sorrendje, ami meghatározza az izotípus váltás lehetőségeit Cm Cd Cg3 Cg1 Ca1 Cg2 Cg4 Ce Ca2 J regions Az ellenanyag izotípus váltás Az immunválasz során egy ellenanyag specificitása (VDJ és VJ) nem változik (az affinitásérés során változhat az affinitása) Az ellenanyagok effektor funkciói az immunválasz során jelentősen változnak Az ellenanyagok képesek a variábilis domén megtartása mellett más konstans régióra váltani, ami más effektor funkciókra teszi képessé az molekulát
C Cδ C3 C1 Cε2 C1 C 2 C4Cε1 C2 C Cδ C Cδ Ig IZOTÍPUSOK Cµ IgM Cγ1 IgG Cγ2 IgG Cγ3 IgG Cγ4 IgG Cα IgA Cε IgE C Cδ C C IgM
CCδ C2 C4 C C Átrendezett DNS IgM-termelő sejt Switch regiók C Cδ, C2, C4 ISOTÍPUS VÁLTÁS Minden izotípus rekombináció produktív Más szignál szekvenciák és enzimek mint a VDJ átrendeződésnél Antigén stimuláció után Nem véletlenszerű Külső jelek irányítják C C Átrendezett DNS IgE-termelő sejt Első RNA átirat C mRNS Hiper IgM szindróma 2. típus Activation Induced Cytidine Deaminase RNS editing enzim NINCS HIPERMUTÁCIÓ ÉS IZOTÍPUS VÁLTÁS nehéz lánc
Cm Cd Cg3 Cg1 Ca1 Cg2 Cg4 Ce Ca2 Sm Sg3 Sg1 Sa1 Sg2 Sg4 Se Sa2 Switch régiók • A C-régiók előtt a DNS-ben ismétlődő szekvenciákból álló „switch” régiók helyezkednek el. (Kivétel a Cd régió) • Az Sm 150 [(GAGCT)n(GGGGGT)] ismétlődő szakaszból áll (n=3 – 7) • Az izotípus váltás mechanikusan sok vonatkozásban hasonló a V(D)J recombinációhoz, DE • Minden rekombinációs esemény produktív • Más rekombinációs szignál szekvenciák és enzimek közvetítik • A B sejt antigén-specifikus aktivációjától függ • Nem véletlenszerű folyamat, mert külső szignálok mint pl. a T sejtek által termelt citokinek befolyásolják • Az izotípus váltás az antigénnel való találkozást követően, az aktivált T-sejtek által termelt citokinek segítségével a perifériás nyirokszervekben megy végbe
Cm Cd Cg3 Cg1 Ca1 Cg2 Cg4 Ce Ca2 Sg3 Cd Cd Cg3 Cm Cm Sg1 Cg3 Cg1 V23D5J4 V23D5J4 Cg3 V23D5J4 Ca1 Ca1 IgG3 termelés IgM IgG3 IgA1 termelés IgM IgA1 IgA1 termelés IgG3 IgA1 V23D5J4 V23D5J4 V23D5J4 Cg3 Ca1 Ca1 Switch rekombináció Minden rekombinációnál a konstans régiók kivágódnak A génsorrend miatt egy IgE – termelő B sejt már nem tud IgM, IgD, IgG1-4 vagy IgA1 termelésre váltani
IMMUNOGLOBULIN OSZTÁLY VÁLTÁS REKOMBINÁCIÓVAL AID (Activation Induced (citidin) Deaminase C →U, RNS editing enzim) elindítja a switch rekombinációt UNG (uracil DNA glycosylase) kivágja az U → bázis mentes vég, AP-endonuleáz/liáz → egyszálú hasítás (nick) Hiba az Ig osztály váltásban - Hiper IgM szindróma, 2 típus, emberben autoszomális
HYPER IgM Szindróma (Autoszómális) - Intrinsic B sejt hiba, az activation induced deaiminase (AID) hiánya okozza, ami a cytidine – uridine reakciót katalizálja - Az enzim szerepet játszik az affinitás érés és az izotípus váltás szabályozásában
VL CDR1 CDR2 CDR3 Complementary Determining Region = hipervariábilis régió
HVR3 150 100 HVR2 Variabilitás Index HVR1 50 FR2 FR1 FR4 FR3 0 25 75 100 50 Amino savak sorszáma N – C terminális A VARIÁBILIS RÉGIÓ SZERKEZETE - VH Hipervariábilis (HVR) vagy komplementaritást meghatározó régiók (CDR) és váz (framework/FR) szakaszok V régió DJ régió Kapcsolási sokféleség
Szomatikus hipermutáció FR1 CDR1 FR2 CDR2 FR3 CDR3 FR4 Variabilitás 100 80 60 40 20 20 40 60 80 100 120 Aminosavszám A különböző specificitású ellenanyagokban található pont mutációk összehasonlítása Wu - Kabat analízissel Mik a következményei az immunválasz során végbemenő mutációknak egy adott epitóp ellen irányuló ellenanyagban? Hogyan befolyásolja az ellenanyag specificitását és affinitását?
SZOMATIKUS HIPERMUTÁCIÓ 0. nap Ag Plazmasejt klónok 1 2 3 4 5 6 7 8 ELSŐDLEGES I.V. AFFINITYMATURATION 9 1011 12 13 14 15 16 14. nap Ag 17 1819 20 21 22 23 24 MÁSODLAGOS I.V.
6. nap 12. nap 8. nap 18. nap Clone 1 Clone 2 Clone 3 Clone 4 Clone 5 Clone 6 Clone 7 Clone 8 Clone 9 Clone 10 Hátrányos mutáció CDR1 CDR2 CDR3 CDR1 CDR2 CDR3 CDR1 CDR2 CDR3 CDR1 CDR2 CDR3 Előnyös mutáció Semleges mutáció A szomatikus hipermutációaffinitás éréshez vezet Kisebb affinitás– Nincs klonális szelekció Nagyobb affinitás– Klonális szelekció Azonos affinitás– Nem hat a klonális delécióra A hipermutációs folyamat aktivált T limfociták közreműködését igényli A mutáció ‘hot spots’ (CDR régiók) körül halmozódnak, amit a kettős láncú töréseket követő hiba javító DNS repair enzim állít helyre
CDR2 H CDR1 CDR3 Antigén determináns CDR1 CDR3 CDR2 L A CDR1 és a CDR2 régiókat a V-gének kódolják A könnyűlánc CDR3 régióját a V és J gének kódolják A nehézlánc CDR3 régióját a V, D és J gének kódolják
A hipervariábilis és váz szekvenciák szerepe • A vázszekvenciák stabilizálják a hipervariábilis hurkokat • A váz kompakt, hidrofób maggal rendelkező hordó alakú blemezes szerkezetet alakít ki • A hipervariábilis hurkok rugalmasabbak mint a blemezek és térszerkezeti egységet alkotnak • A hipervariábilis hurkok szekvenciája a különböző specificitású ellenanyagok között nagyon sokféle • A hipervariábilis hurkok aminosav szekvenciája meghatározza az ellenanyag kötőhely alakját, hidrofób jellegét és töltés viszonyait • A hipervariábilis hurkok sokfélesége biztosítja a sokféle antigénnel reagálni képes ellenanyag készlet sokféleségét
B-SEJT AKTIVÁCIÓ (HOL ÉS HOGYAN TÖRTÉNIK?)
A B-sejt aktiváció fő lépései AKTIVÁCIÓ PROLIFERÁCIÓ/DIFFERENCIÁCIÓ FELISMERÉS Ea termelés Klonális szaporodás Segítő T Izotípus váltás Affinitás érés Memória B sejt
B sejt a vérben T sejtesterület B sejtesterület Efferens/kivezető nyirokér A B-sejtek recirkulációja antigén hiányában (nyirokcsomó)
B sejtek gyors prolife- rációja B sejtek a HEV-en keresztül a nyirokcsomóba jutnak Antigen az afferens/bevezető nyirokéren át a nyirokcsomóba jut Y Y Y Y Y Y Y Y Y Y Y Y Y Y B sejtek elhagyják a germinális centrumot és plazmasejtté differenciálódnak Y Y Y Y GERMINÁLIS CENTRUM Átmeneti struktúra, gyors proliferáció A keringő B sejteket az antigének tartják vissza a limfoid szervekben
„Nagy találkozás” a perifériás nyirokszervekben Az antigént kötő B sejtek a T-sejt területen akadnak fenn Az antigént kötő B sejtek az aktivált T-sejtekkel lépnek kapcsolatba
A germinális centrum szerkezete Szomatikus hypermutáció LZ FDC DZ Szomatikus hypermutáció LZ: világos zóna DZ: sötét zóna FDC: follikuláris dendritikus sejt
Az antigén a follikuláris dendritikus sejtek (FDC) felszínéhez kötődik (FDC) Az FDC-ek immun komplexeket kötnek (Ag-Ab) Az antigén ingert követően 12 hónappal is kimutatható az FDC-ek felszínén Egyetlen sejt sokféle antigént köthet Az FDC felszínén az immunkomplexek un. iccosomákat képeznek, melyek leválhatnak és utána a környező GC B sejtek felveszik őket A B sejtek az FDC felszínén ismerik fel az antigént
T SEJT FÜGGŐ B SEJT AKTIVÁCIÓ A LIMFOD SZERVEKBEN IgM Naiv B sejt Nem mutált plazma sejt Antigén Th sejt Szomatikusan mutált plasma sejt IgG IgA IgE B sejt blaszt Csíraközpont B sejt Memória B sejt
Ig domain + CHO a b ITAM ITAM Y Y Y Y SIGNALING UNITS OF THE B-CELL RECEPTOR Ig-a/CD79a Ig-b/CD79b ITAM: YxxLx7YxxI ITAM: Immunoreceptor Tyrosine-based Activation Motif
A szignál transzdukció lépései B-sejtekben Membrán Ig keresztkötése Ag által Tirozin foszforiláció Biokémiai intermedierek Transzkripciós faktorok Aktív enzimek
A B-sejt receptor keresztkötés következményei Ag kötés, a felszíni Ig keresztkötése Limfocita aktiváció Fenotípus/funkció változás Fokozott túlélés Osztódás Ko-stimuláló molekulák kifejeződése Citokin receptorok kifejeződése Kivándorlás a limfoid tüszőből a T sejtes területekre
A LIMFOCITA AKTIVÁLÁS KINETIKÁJA, KÖVETKEZMÉNYEI Nyugvó limfocita G0 Ko-receptor Adhéziós molekula Citokinek 2. JEL Effektor sejtMemória sejt Transzport folyamatok Membrán változás RNS és fehérje szintézis sejtosztódás DNS szintézis Limfoblaszt PTK aktiváció RNS szintézis Szabad Ca++ Fehérje szintézis Fehérje foszforiláció DNS szintézis Nyugvó limfocita G0 0 10sec 1min 5min 1óra 6 óra 12 óra 24 óra ANTIGÉN 1. JEL
A CR2 (CD21) KOMPLEMENT RECEPTOR A B – LIMFOCITÁKON KO-STIMULÁLÓ SZEREPET TÖLT BE C3d ANTIGÉN Antigén determináns CR2/CD21 CD19 B-SEJT TAPA=CD81 Y Y Fokozott B-sejt aktiválás
A B-SEJT RECEPTORON ÁT KÖZVETÍTETT JELET A CD22 NEURAMINSAV RECEPTOR GÁTOLJA Mannóz Testi sejtek Baktérium Antigén B-SEJT Sziálsav, glikokonjugátumok, T sejt CD45 CD22 ITIM/ITAM
Ellenanyagokkal és komplement faktorokkal opszonizált Epstein Barr Virus (EBV) elektromikrográfiás képe Negativ festésű EBV Ellenanyagokkal és komplement komponensekkel körbevett EBV Ellenanyagokkal körbevett EBV
A KÖTŐHELY SAJÁTSÁGAI MÉRET ALAK HIDROFÓB HIDROFIL POSITÍV TÖLTÉSŰ NEGATÍV TÖLTÉSŰ AZ ANTIGÉN ÉS ELLENANYAG KAPCSOLÓDÁST NEM-KOVALENS KÖLCSÖNHATÁSOK BIZTOSÍTJÁK Egy kötőhely több antigént is köthet A kötés affinitása/aviditása széles tartományban változik
Növekedési faktorok Adhéziós molekulák MHC – peptid - TCR Adhéziós molekulák ELLENANYAGOK Kötés erősség
A B – SEJT DIFFERENCIÁCIÓT A T-SEJTEK SEGÍTIK ANTIGÉN CITOKINEK PLAZMA SEJT B-SEJT IZOTÍPUS VÁLTÁS ÉS AFFINITÁS ÉRÉS CSAK T-SEJT SEGÍTSÉGGEL MEGY VÉGBE HOGYAN LÁTJÁK A T-SEJTEK AZ ANTIGÉNT?