Download
1 / 28
290 likes | 499 Views
Nightjars, owlet-nightjars, potooes, oilbird, frogmouths, hummingbirds, swifts. Heterogeneous assemblage of enigmas. Accipitrid diurnal raptors, osprey and secretarybird, rollers, woodpeckers, trogons, mousebirds, owls. Parrots and Passerines.

E N D
Nightjars, owlet-nightjars, potooes, oilbird, frogmouths, hummingbirds, swifts Heterogeneous assemblage of enigmas Accipitrid diurnal raptors, osprey and secretarybird, rollers, woodpeckers, trogons, mousebirds, owls Parrots and Passerines Pelicans, cormorants, herons, storks, cranes, rails, loons, penguins, albatrosses, cuckoos, turacos, bustards Shorebirds, gulls, auks Pheasants, quails, currasows, ducks, geese, swans Ratites and Tinamous Ericson 2008
Nightjars, owlet-nightjars, potooes, oilbird, frogmouths, hummingbirds, swifts Heterogeneous assemblage of enigmas Suboscine (represented by Tyrannida) Oscine (represented by Passerida) PASSERINES Largest Order of Extant Birds Accipitrid diurnal raptors, osprey and secretarybird, rollers, woodpeckers, trogons, mousebirds, owls Parrots and Passerines Pelicans, cormorants, herons, storks, cranes, rails, loons, penguins, albatrosses, cuckoos, turacos, bustards Shorebirds, gulls, auks Pheasants, quails, currasows, ducks, geese, swans Ratites and Tinamous Ericson 2008
Timing Red = Australasia Green = Africa and Eurasia Blue = North and South America Grey = ambiguous ancestral / current area (Barker et al. 2004)
Routes (Ericson et al. 2002) (Cracraft 2001)
Syrinx Gill 2005
Morphology of Voice Brackenbury 1982 and Gill 2005
Syrinx of Suboscine and Oscine Gill 2005 and Wallace and Mahan 1975
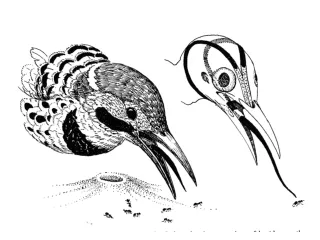
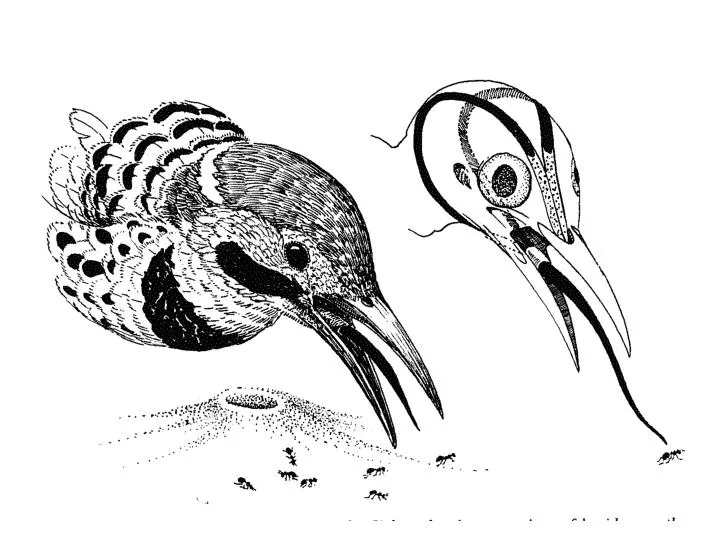
Rictal Bristles • Tyrannidae
Bird Brains? • Transitive inference • Episodic memory • Object constancy (follow disappeared object) • Tool manufacture • Social learning • Theory of mind (Nihei and Higuchi 2001)
All Bird Brains are Not Equal (Emery and Clayton 2004)
Hippocampus • In birds and mammals most medial part of pallium • Spatial memory (caches, location of danger) • Changes in size with seasonal needs
Lateralization and Brain Complexity • Complex neural connections and lateralization • Left hemisphere for complex integration and learning • New Caledonian Crows are mostly right-billed (tilting to use right eye), using left hemisphere to guide tool making and use • Song learning is also controlled from left hemisphere (Cnotka et al. 2008)
Scrub-jays Florida Scrub-jay Fig. 1. FROM DELANEY et al. 2008---Distribution of Island and Western scrub-jays, with associated geographic trends in morphological characteristics. Species distributions are adapted from Curry et al. (2002); orange = Aphelocoma insularis, green = Californica group of Aphelocoma californica, light blue = Woodhouseii group of Aphelocoma californica, and dark blue = sumichrasti group of Aphelocoma californica. Currently we recognize 3 species, but there are most likely 5 and maybe 6 isolation has been of paramount importance and novel selective pressures from foods eaten (oaks versus other seeds)
50ka During the last several million years land connections (via Beringia) between New and Old Worlds waxed and waned with glaciation. Beringia was dry and offered land passage.
Corvids Invade Our World Tied to changes in vegetation and sea level Tertiary forests of Australia were being replaced by deserts perhaps forcing corvid ancestors (related to Birds of Paradise and Orioles) to leave Australia and head north in Oligocene and Miocene to Asia, following northward movement of tropical forests 6-8 mya in Miocene New World Jay ancestor from forests of southeast Asia, radiate in South America (first) and North America 15,000 ya – 2 mya in Pleistocene Old World Jay (Gray Jay) Nutcracker Magpie Crow Raven (from article on evolution of cats; Johnson et al. 2006; Science 311:73-77)
Ravens • 4 clades diverging in Africa 1.7-3.8my • Corvus corax ancestor diverges (closest relative is C. albus) shortly thereafter • C. corax invades New World 2my and new and old world ravens begin independent evolution • Old world raven spins off Canary Island Raven 650,000 yr • New world ravens spins off Chihuahuan Raven • C. corax reinvades New World 15,000 years ago
Complexity Revealed By Genetic Analyses • Common Raven • ~15,000 years ago old world ravens again invaded the new world via Beringia • Holarctic and California clade of ravens now found in North America, but they are becoming more similar, not diverging as they had in past. • Giving us new insights into what constitutes a “species”
Wrens • New World Family Winter Wren Vocalizations
Bushtits Male Female
Mixed Species Flocks • Nonbreeding season • Kinglets, chickadees, woodpeckers, creepers, nuthatches • Tropical tanagers, euphonias, toucans, woodpeckers, and lots more • Predator avoidance, food finding
Importance of Snags and Woodpeckers to Secondary Cavity Nesters • Creepers, Nuthatches, Chickadees, Swallows
Habitat Losses • Shrub Steppe, Sagebrush, Thornscrub • Shrikes, grouse, many others • Grasslands • larks
Literature • Barker, F. K., Cibois, A., Schikler, P., Feinstein, J., and J. Cracraft. 2004. Phylogeny and diversification of the largest avian radiation. Proc. Natl. Acad. Science 101:11040-11045. • Butler, A. B. and R. M. J. Cotterill. 2006. Mammalian and avian neuroanatomy and the question of consciousness in birds. Biological Bulletin 211:106-127. • Cnotka, J., Gunturkun, O., Rehkamper, G., Gray, R. D. and G. R. Hunt. 2008. Extraordinary large brains in tool-using New Caledonian crows (Corvus moneduloides). Neuroscience Letters 433:241-245. • Cracraft, J. 2001. Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc. Royal Soc. B. 268:459-469. • Delaney, K. S., Zafar, S., and R. K. Wayne. 2008. Genetic divergence and differentiation within the western scrub-jay (Aphelocoma californica). The Auk 125:839-849. • Emery, N. J. 2006. Cognitive ornithology: the evolution of avian intelligence. Phil. Trans. R. Soc. B 361:23-43. • Emery, N. J. and N. S. Clayton. 2004. The mentality of crows: convergent evolution of intelligence in corvids and apes. Science 306:1903-1907. • Ericson, P. G. P. 2008. Current perspectives on the evolution of birds. Contributions to Zoology 77:109-116. • Ericson, P.G.P., Anderson, C.L., Britton, T., Elzanowski, A., Johansson, U.S., Kallersjo, M., Ohlson, J.I., Parsons, T. J., Zuccon, D., and Mayr. Gl. 2006. Diversification of Neoaves: integratino of molecular sequence data and fossils. Biology Letters 2:543-547. • Ericson, P. G. P., Christidis, L., Cooper, A., Irestedt, M., Jackson, J., Johansson, U. S., and J. A. Norman. 2002. A Gondwanan origin of passerine birds supported by DNA sequences of the endemic New Zealand wrens. Proc. Royal Soc. B. 269:235-241. • Ericson, P. G. P., Jansén, A-L, Johansson, U. S., and J. Ekman. 2005. Inter-generic relationships of the crows, jays, magpies and allied groups (Aves:Corvidae) based on nucleotide sequence data. Journal of Avian Biology 36:222-234. • Omland, K. E., C. L .Tarr, W. I Boarman, J. M. Marzluff, and R. C. Fleischer. 2000. Cryptic genetic variation and paraphyly in ravens. Proceedings of the Royal Society of London B. 267:2475-2482. • Omland, K. E., J. M. Baker, and J. L. Peters. 2006. Genetic signatures of intermediate divergence: population history of Old and New World Holarctic ravens (Corvus corax). Molecular Ecology 15:795-808. • Peterson, A. T. 1993. Adaptive geographic variation in bill shape of Scrub Jays (Aphelocoma coerulescens). American Naturalist 142:508-527

















![[PDF] EZ Battery Reconditioning™ by Tom Ericson](https://cdn4.slideserve.com/8498390/ez-battery-reconditioning-by-tom-ericson-dt.jpg)