Download
1 / 55
550 likes | 765 Views
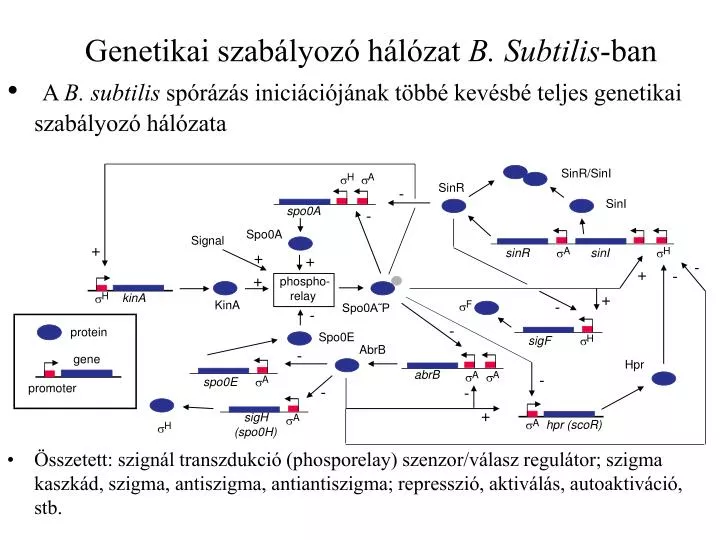
SinR/SinI. SinR. -. SinI. spo0A. -. protein. Spo0A. Signal. gene. +. H. H. A. H. A. A. A. H. F. A. A. H. A. . . . . . . . . . . . . . sinR. sinI. +. +. -. promoter. +. -. +. phospho- relay. +. kinA. spo0E. -. KinA. Spo0A˜P. -.

E N D
SinR/SinI SinR - SinI spo0A - protein Spo0A Signal gene + H H A H A A A H F A A H A sinR sinI + + - promoter + - + phospho- relay + kinA spo0E - KinA Spo0A˜P - - Spo0E sigF AbrB - Hpr abrB - - - + sigH (spo0H) hpr (scoR) Genetikai szabályozó hálózatB. Subtilis-ban • A B. subtilis spórázás iniciációjának többé kevésbé teljes genetikai szabályozó hálózata • Összetett: szignál transzdukció (phosporelay) szenzor/válasz regulátor; szigma kaszkád, szigma, antiszigma, antiantiszigma; represszió, aktiválás, autoaktiváció, stb.
TÍPUS HELY TERMÉK RELATÍV AKTIVITÁS a AMANITIN ÉRZÉKENYSÉG POL I NUCLEOLOUS rRNS 50-70 % - POL II NUCLEOPLASMA mRNS 20-40 % + POL III NUCLEOPLASMA kis RNS (tRNS & 5S rRNS) 10 % VÁLTOZÓ EUKARIÓTA RNS POLIMERÁZOK
L’ L L’ L L’ L RNS POLIMERÁZOK ÖSSZEHASONLÍTÁSA EUKARIÓTA E. coli a2bb’ I II III ~ bésb’ b b’ CTD ~ aésa’ a a’ KÖZÖS
A génexpresszió szabályozása eukariótákban: • Rövid távú, • hosszú távú – génszabályozás: fejlődés/differenciálódás A Drosophilaegyedfejlődés szabályozása
Az eukarióta gének is fehérje-kódoló egységekként és a szomszédos szabályozó régiókkal szabályozódnak, de nem operonok. • Az eukarióta gének szabályozása komplexebb, mert van sejtmagjuk. • (a transzkripció és a transzláció elkülönülten). • Az eukarióta gének szabályozásának két kategóriája: • Rövid távú – gének gyors ki/bekapcsolása a környezetre és a sejt igényeire adott válaszként. • Hosszú távú – az egyedfejlődés és a differenciálódás génjei.
Az eukarióta gének hat szinten szabályozódnak: transzkripció RNS processzálás mRNStranszport mRNS transzláció mRNS degradáció protein degradáció
A transzkripció szabályozása: • Promóterekkel • A transzkripciós starthely előtt. • Néhány azt határozza meg, hol kezdődjön a transzkripció (pl. TATA), míg mások, hogy megkezdődjön-e. • A promóterek aktivációját specifikus transzkripciós faktor (TF) proteinek (specifikus TF-ekkapcsolódnak specifikus promóterekhez). • Egy adott génnek egy, vagy több promótere (mindegyik specifikus TF fehérjékkel) lehet. • A promóterek lehetnek pozitívan és negatívan szabályozottak.
Enhancerekkel A transzkripciós starthely előtt, vagy után vannak. • A szabályozó fehérjék specifikus enhancer szekvenciákhoz kötődnek; a kötödést a DNS szekvencia határozza meg. • A TF-ekhez kötődő DNS hurkot képez és így teremti meg a kapcsolatot az enhancer elemekkel. • A szabályozó fehérjék közötti kölcsönhatások határozzák meg, hogy a transzkripció aktiválódik,vagy represszálódik, azaz pozitívan, vagy negatívan szabályozott lesz-e. • Például
Még a promóterekről és az enhancerekről: • Néhány szabályozó fehérje minden sejttípusban van, mások specifikusak. • Minden promóternek és enhancernek az expressziót meghatározó specifikus fehérje készlete van. • A génmegnyilvánulás mértékét a pozitívan és a negatívan szabályozó fehérjék közötti kölcsönhatások alakítják. • Kombinatorikus génszabályozás; enhancerekés promóterek sok hasonló szabályozó fehérjét tudnak kötni, számtalan kölcsönhatási lehetőséggel.
Kromoszómaszerkezet, az eukarióta kromoszóma hisztonokkal van csomagolva: • A prokariótáknak nincsenek hisztonjaik és más struktúr fehérjéik, így a így a DNS közvetlenül elérhető. • Az eukariótáknak vannak hisztonjaik és a hisztonok represszálják a transzkripiót, mert interferálnak a DNS-kötő fehérjékkel. • Igazolás DNáz I érzékenységi kísérletekkel: • A DNáz I lebontja a transzkripciósan aktív DNS-t. • A nem-átíródó DNS-t a hiszton pajzs megvédi a DNáztól és nem tudja olyan könnyedén lebontani. • Ha kísérletesen adunk hisztonokat és és promóter-kötő fehérjéket, a hisztonok kompetitíven kapcsolódnak a promóterhez és gátolják a transzkripciót. • Megoldás: a transzkripciósan aktív géneknek lazább a kromoszóma szerkezete, mint az inaktív géneké. • A hisztonok acetilezése és foszforilezése megváltoztatja DNS-kötő képességüket. • Az enhancer-kötő fehérjék kompetitíven blokkolják a hisztonokat, ha kísérletesen együtt adjuk hisztonokkal és és promóter-kötő TF-ekkel. • Az RNS polimeráz és a TF-ek körülfogják a hiszton/nukleoszómát és lesz transzkripció.
DNS metiláció és a transzkripció: • Az újonnan szintetizálódott DNS kis mennyisége (~3% emlősökben) metilezéssel kémiailag módosított. • A metiláció leggyakrabban szimmetrikus CG szekvenciáknál. • A transzkripciósan aktív gének szignifikánsan kisebb mértékben metilezettek, mint az inaktív gének. • A metiláció génje esszenciális az egér egyedfejlődésénél (egy gén kikapcsolása szintén lehet fontos). • A törékeny X szindróma emberi betegséget is metilezés eredményezi; az FMR-1 géntmetilezés csendesíti.
Rövid távú transzkripció szabályozás – a galaktóz hasznosítás génjei élesztőben: • 3 gén (GAL1, GAL7, & GAL 10) kódolja a galaktóz metabolizmus útvonalban működő enzimeket. • GAL1 galaktokináz • GAL7 galaktóz transzferáz • GAL10 galaktóz epimeráz • Az útvonalvégén D-glükóz 6-foszfát képződik, ami belép a glikolízis útvolalba és folyamatosan átíródó gének metabolizálják. • Galaktóz nélkül, aGAL gének csendesek. • A GAL gének glükóz hiányában és galaktóz jelenlétében gyorsan indukálódnak. • Analógia: azE. colilac operon repressziójaglükózzal.
A GAL gének egymáshoz közel, de nem egy operonban vannak. • Egy másik nem-kapcsolt gén, aGAL4, egy represszor fehérjét kódol, ami egy promóter elemhez kötődik:upstream aktivátor szekvencia (UASG). • Az UASGaGAL1és aGAL10között helyezkedik el. • Transzkripció azUASG-től mindkét irányba. • Ha nincs galaktóz, aGAL4terméke (GAL4p) és egy másik géntermék (GAL80p) kötődik azUASGszekvenciához; nem lesz transzkripció. • Ha galaktózt adunk, a galaktóz metabolit kapcsolódika GAL80p-velés aGAL4paminosavak foszforileződnek. • A galaktóz inducerként működik, konformáció változást okoz a GAL4p/GAL80p-ben.
Hormonális szabályozás – egy másik jó példa a rövid távú transzkripció-szabályozásra: • A magasabb rendű eukarióták sejtjei specializálódottak és általában jól védettek a külső környezet gyors változásaitól. • A hormon szignálok az egyik mechanizmus, amivel a környezetre reagálva szabályozzák a transzkripciót. • Az egyik sejt által termelt hormonok inducerként hatnak és fiziológiai választ váltanak ki egy másik sejtben. • A hormonok csak a hormon-specifikus receptorokkal rendelkező célsejtekre hatnak és a hormonszintet feedback útvonalak állítják be. • A hormonok két módon szolgáltatnak jelet: • A szteroid hormonokátmennek a membránon és a citoplazmás receptorokhoz kapcsolódnak, együtt kötődnek a DNS-hez és szabályozzák a génexpressziót. • A polipeptid hormonoka sejtfelszíni receptorokhoz kapcsolódnak és aktiválják a transzmembrán enzimeket, hogy másodlagos hírvivőket szintetizáljanak (pl. cAMP), amelyek a aktiválják a transzkripciót.
Emlős szteroid hormonok. Növény hormonok
Hormonális szabályozás: • A szteroid hormonokkal regulált gének szekvenciáiban van kötő régió: szteroid hormone response elements (HREs). • HRE-k gyakran több kópiában is az enhancer szekvencia régióban. • Ha nincs szteroid: • a receptor beborítvaésdajka fehérjék (chaperone) „vigyázzák”; nincs transzkripció • Ha van szteroid: • A szteroid lecseréli a dajka fehérjéket, kötődik a receptorhoz, majd a HRE szekvenciához; megkezdődik a transzkripció.
Szabályozás RNS processzálódással: • Az RNS processzálódás szabályozza az mRNS képződését a prekurzor RNS-ekből. • Két független szabályozó mechanizmus van: • Alternatív poladeniláció = amikor a polyA farok szintetizálódik • Alternatív splicing = amikor az exonokvágódnak ki • Az alternatív pliadeniláció és splicing együtt is előfordulhatnak. • példák: • Humán kalcitonin (CALC) géna thyroid (pajzsmirigy)és a neuron sejtekben • Ivar meghatározásDrosophila-ában
Az ivar meghatározása Drosophila-ban alternatív splicing-gal. • Az ivart az X:A arány határozza meg. (X=kromoszóma; A=allél) • Sxl (sex lethal) génhatározza meg az utat a hímeknek és a nőstényeknek. • Ha X:A = 1, az összes intron és az exon 3 (ami stop kodont tartalmaz) is kivágódik. • Ha X:A = 0.5, nem képződik funkcionális fehérje. Átíródik, jó splicing Átíródik, rossz splicing Jó Sxlp Rossz fehérje Jó tra splicing Rossz alternatív Trap- Trap+ Trap+Tra-2p női dsx splicing Dsx fiú splicing Fiú diff. gátolt Leány diff.
mRNS transzport szabályozás: • Az eukarióta mRNS transzport szabályozott. • Néhány kísérlet azt mutatta, hogy az elsődleges transzkriptimok ~1/2-e soha nem hagyja el a sejtmagot és lebontódik. • Az érett mRNS-eka nukleáris pórusokon keresztül távoznak.
mRNS transzlációs szabályozás: • Például a megtermékenyítetlen petébenaz mRNS-ek (a petében tárolt /nincs új mRNS szintézis) transzlációja a megtermékenyítés után azonnal megindul. • A tárolt mRNS-eket védő fehérjék akadályozzák a transzlációt. • Poli(A) farok segíti a transzlációt. • A tárolt mRNS-eknek általában rövid poli(A) farka van. (15-90 A vs 100-300 A). • Specifikus mRNS-ek kijelölődnek a dezadenilációra (“tail-chopping”) a betárolás előtt az AU-gazdag 3’-UTR szekvenciáik alapján. • Akkor aktiválódnak, ha egy enzim felismeri az AU-gazdag szekvenciát és még hozzáad ~150 A-t és a ploi(A) farok eléri teljes hosszúságát.
mRNS degradációs szabályozás: • A citoplazmában minden mRNS ki van téve degradációnak. • A tRNS-ekés az rRNS-ek általában nagyon stabilak; mRNS-ekstabilitása változó (percek, hónapok). • A stabilitás a szabályozó szignálok hatására változhat és úgy vélik, hogy ez a legfontosabb szabályozó pont. • Különböző szekvenciák és folyamatok befolyásolják az mRNS félélet-idejét: • AU-gazdag elemek • Másodlagos szerkezet • Dezadenilációs enzimek • 5’ de-capping • Vágás az mRNS belsejében és a fragmentumok degradációja.
Poszt-transzlációs szabályozás - protein degradáció: • A fehérjék lehetnekrövid-(pl. szteroid receptorok) vagyhosszú-életűek (pl. a szemlencse fehérjéi). • A fehérjék degradációjához az eukariótákban szükséges az ubiquitin kofaktor. Az ubiquitin kötődik a fehérjékhez és megjelöli azokat a proteolítikus enzimeknek. • Az N-terminális aminosavtól függ a fehérje stabilitása és meghatározza az ubiquitin kapcsolódás mértékét. • Arg, Lys, Phe, Leu, Trp 1/2 élet ≤3 perc • Cys, Ala, Ser, Thr, Gly, Val, Pro, Met1/2 élet ≥ 20 óra
Hosszú távú génszabályozás - fejlődés és differenciálódás: Egy adott genotípusú, egyetlen egy sejtből számos specializált szövet és szerv képződik. Fejlődés A fejlődés a genom kölcsönhatása a citoplazmával és a külső környezettel, hogy egy programozott lépéssorral jellemző irreverzibilis események jöjjenek létre. Differenciálódás: Differenciálódással sejttípusok, szövetek és szervek jönnek létre specifikus génszabályozással. A fejlődés és a differenciálódás több szinten tanulmányozható: • Morfológia • Biokémia • Aktivációs-repressziós mintázat
Genom aktivitás • Az eukarióta DNS nagy része nem kódol fehérjét. • Tengeri sün, az egyedi szekvenciák ~6%-a aktív egyszerre egy időben. • Az érett szövetekben az expresszió ~0,8%-ra csökken. • A nem-átíródó DNS legnagyobb részének funkciója nem ismert.
A DNS állandósága a fejlődés alatt • A felnőtt sejtjeinek ugyanolyan a genomja, mint a zigótának? • Frederick Steward (1950) sikeresen növesztett új sárgarépát a növény szárszövetéből. • Ian Wilmut és mtsi (1997) birkát klónoztak. • 1997 óta egeret, marhát, majmot és humán embriósejtet klónoztak. • A differenciálódott szomatikus sejt nukleusza totipotens; mindazzal a genetikai információval rendelkezik, ami a fejlődéshez szükséges.
A fejlődés és a differenciálódás modell szervezetei: • Drosophila melanogaster (ecet muslica) • Régóta vizsgált. • Sok, a fejlődést befolyásoló mutáció azonosítottak. • Caenorhabditis elegans (nematóda, fonalféreg) • Kis genom (97 Mb) • A testi sejtek száma és a leszármazása ismert. • A genetikai keresztezés és az öntermékenyítés könnyű. • A test áttetsző. • Brachydanio rerio (zebradánió) • Az embriók áttetszőek. • Nagyszámú hal tenyészthető. • Screening technikák jól kidolgozottak. • Arabidopsis thaliana (növény, libatop) • Kicsi, könnyű keresztezni és elemezni nagyszámú utódot. • Sok fejlődési mutánst azonosítottak.
ADrosophilafejlődés genetikai szabályozása: ADrosophilafejlődés stádiumai: (10-12 nap) Tojás Lárva (3 instars) Báb Felnőtt
ADrosophilaembrió fejlődése: • A fejlődés a megtermékenyítéssel kezdődik. • A megtermékenyítés előtt, a petében molekuláris gradiens van. Poláriscitoplazmaalakul ki a posterior végén. • A 2 nukleusz fuzionál a megtermékenyítés után és zigóta képződik. • 9 mitotikus osztódás lesz sejtosztódás nélkül és 7 osztódás után néhány nukleusz a poláris citoplazmához vándorol (posterior) és kialakítja a csíra-vonal prekurzorokat. • Más nukleuszok a sejtfelülethez vándorolnak és kialakul a blastoderma prekurzor. • 4 további mitotikus osztódás az összes sejtmag elkülönül egymástól sejtmembránnal.
A fejlődés egymást követő lépései két folyamattól függenek: • Az anterior-posterior és a dorsal-ventral molekuláris gradiensektől . • A (1) paraszegmentumokés az (2)embrionális szegmentumok képződése, amelyek végül a (3) felnőtt szegmentumok képződéséhez vezetnek. A felnőtt szelvényezettsége az embrióénak felel meg.
A gének három fő csoportja szabályozza a fejlődést és a differenciálódást *Letális és abnormális szerkezetek a fejlődés közben. Anyai hatású gének Szegmentáció gének Homeotikus gének
1. Anyai hatású gének (az anya készítette elő) bicoid gén • Az anterior szerkezet kialakulását szabályozza (a mutánsoknak mindkét végükön posterior szerkezet van). • A gén a peteképződés közben íródik át és a megtermékenyítés után nyilvánul meg. • A bicoid géntermék fehérje gradienst alakít ki. nanos gén • A potroh képződést szabályozza (mRNS-ek a pete posteriorrészén gyűlnek össze). • A nanos gén szintén fehérje gradienst alakít ki. torso gén • Transzkripció és transzláció a peteképződés közben. • A petében mindenhol van, de csak a plólusokon aktív.
2. Szelvényezettség gének (az embriót régiókra osztja): ~25 szegmentációs gént azonosítottak és osztottak 3 csoportba: • Gap gének – a mutációjuk eredményeként néhány szomszédos szelvény deléciója. • Pair rule gének – mutációjuk minden szegmensben a mintázat ugyanazon részének delécióját okozza. • Szegmens polaritás gének – mutánsoknál a szegmens egy része kicserélődik a kapcsolódó fél-szegmens tükörképével.
A szelvényezettség gének szerepe mutánsokkal meghatározva
3. Homeotikus gének (a szerkezetet határozzák meg): • A homeotikus gének minden egyes szelvénynél meghatározzák a testrész fejlődését. • A felnőtt testrészei nem-differenciálódott lárva szövetekből, az imágó diszkekből fejlődnek ki. • A homeotikus mutánsok más testrészt növesztenek, mint a normális testrész egy bizonyos szelvényen. • A különböző homeotikus géncsoportoknak van azonos szekvenciája ~180 bp, a homeoboxok, amik fehérjét kódolnak. • A homeoboxok szabályozzák a fejlődést és a termelt fehérjék a gén egységek upstream régiójához kötődnek. • A homeotikus génkomplexeketHox-nak rövidítjük. • Hox génekgerincesekben és növényekben is.
A homológ imágó diszkek elhelyezkedése a lárvában és felnőttben.
Homeotikus mutánsok bithorax homeotikus mutáns antennapedia 4-szem
Abithorax homeotikus gének szerveződéseDrosophila-ában, ~300 kb.