Download
1 / 58
580 likes | 837 Views
Horizontální genový přenos. Genomy v pohybu. Vertikální genový transfer – přenos genetického materiálu z rodičů na potomky (replikace DNA) Horizontální (laterální) genový transfer (HGT, LGT) – přenos genetického materiálu mezi druhy. Hnací síly evoluce genomů Rekombinace (sex, meióza)

E N D
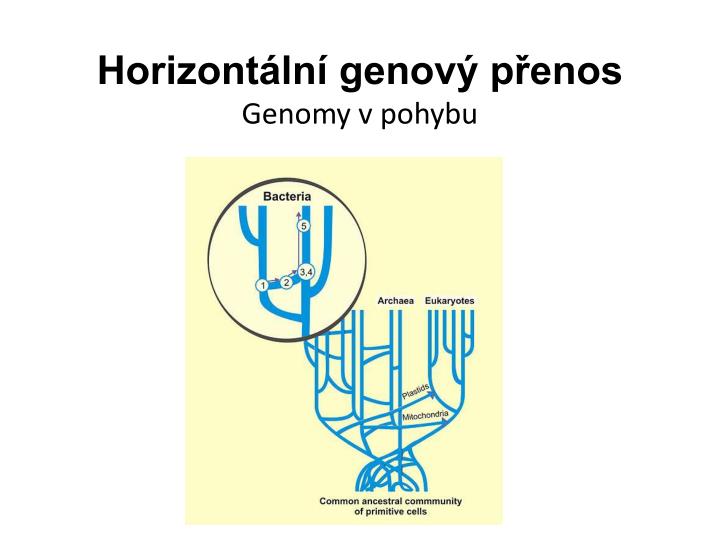
Horizontální genový přenos Genomy v pohybu
Vertikální genový transfer – přenos genetického materiálu z rodičů na potomky (replikace DNA) • Horizontální (laterální) genový transfer (HGT, LGT) – přenos genetického materiálu mezi druhy
Hnací síly evoluce genomů • Rekombinace (sex, meióza) • Bodové mutace (pomalé) • HGT (zdroj rychlých adaptací)
Význam HGT • Nahrazení genů pro existující dráhy • Přínos nové funkce (metabolické, inovace, adaptace) • Evoluce organel Encephalitozoon cuniculi Pyrenophora tritici-repentis ←Stagonospora nodurum
Rozsah HGT • 20% genomu bakterií • < 1% genomu protist • 5% u extremofilní červené řasy Galdieria sulphuraria • 4% genomu u bachorových nálevníků
HGT v evoluci metabolizmu baktérií • Rezistence na antibiotika a toxické kovy • Fotosyntéza • Aerobní respirace • Fixace dusíka, redukce síry, syntéza izoprenoidů • Tolerance extrémních podmínek (vysoké teploty, salinita) • Bioremediace (přeměna kontaminací pomocí mikroorganismů) • Atd.
Mechanizmy HGT Baktérie: konjugace, transformace, transdukce Eukaryota: neznáme (zřejmě potrava a viry)
Mobilní genetické elementy • Mobilom – souhrn mobilních genetických elementů, které jsou schopné přesunu uvnitř genomu i mezi genomy • Transposony (rozšířené u eukaryot) • Plazmidy • Bakteriofágy (u bakterií) • Molekuly DNA schopné samosestřihu
Transposony („jumping genes“) • Transpozice • Funkce promotorů, zesilovačů, tlumičů transkripce, místa epigenetických modifikací a alternativního sestřihu • Citlivá místa pro rekombinaci → delece, duplikace, inverze, translokace • Vliv na velikosti genomů (C-value) Carlton JM. Science. 2007;315(5809):207-12.
Plasmidy • Extra-chromozomální DNA schopná autonomní replikace • Přirozený výskyt u bakterií (plasmidy virulence), občas u eukaryot (např. S. cerevisiae, E. histolytica) • High copy (>100 kopií) x low copy (1-25 kopií); 1 – 400 kbp • Šíření pomocí konjugace (spojení nepříbuzných organizmů)
Bakteriofágy (viry) • Nejrozšířenější entity v biosféře • Regulují populace bakterií • Hlavní přispěvatelé procesu HGT (životní cyklus fága) • Přínos nových funkcí
Mechanizmus HGT u eukaryot • Fagotrofie – hypotéza „You are what you eat“ (Doolittle W.F, 1998) • Virální transdukce - nucleocytoplasmic large DNA viruses („gene pickpockets“) u protist a řas • Jiný – konjugace bakterií a hub anebo transformace volnou DNA z prostředí
Jak odhalit HGT? • Nesoulad fylogeneze genu s fylogenezí organismu • Nerovnoměrná distribuce genu („patchy distribution“; přítomnost/absence genu) • Anomálie ve stuktuře DNA • Optimální je kombinace všech
Jak odhalit HGT? Metody molekulární fylogenetiky a molekulární biologie Fig. 28.5. Some trees show unusual relationships for eukaryotes that are not easy to explain within the framework of the current consensus for eukaryotic phylogeny. Entamoeba and the diatom Thalassiosira are closely related in the tree for glutamine synthase (EC 6.3.1.2), but no external data suggest that these two eukaryotes are closely related. One possible explanation is eukaryote-to-eukaryote HGT. Trichomonas vaginalis also contains a homologue of glutamine synthase, but in this case it clusters weakly with Fusobacterium. Alsmark UC et al. 2009
Jak odhalit HGT? • Odlišný poměr GC párů v DNA (přednostně se ale přenáší geny s podobným GC poměrem jak má genom recipienta) • Hustota genů a poměr mobilních elementů Příklad: u zelené řasy Ostreococcus tauri pochází celý chromozom 19 z externího zdroje Derelle et al. 2006 (PNAS)
Hypotéza komplexity • HGT se týká především genů operačních (metabolických) • Informační geny(pro transkripci a translaci) jsou HGT postižené zřídka (informační geny jsou typicky součástí komplexních systémů, jejich přenos by byl složitý. Např. ribozom). • Výjimky např. EF-1α x EFL; α-tubulin u Andalucia; aminoacyl- tRNA syntetázy
Jste to, co jíte, na čem žijete, co žije na vás a ve vás • Životní styl organismů urychluje horizontální přenos genů (fagotrofie, symbiózy): • A.Fagotrofie - pohlcování bakterií anebo menších eukaryot (např. nálevníci Didinium vs. Paramecium) zlepšuje přístup organismu k cizorodé DNA • Těsný kontakt jader u Kryptoperidinium (obrněnky a rozsivky) • Zelené plastidy z pohlcené kořisti (zelená řasa) uvnitř mořského slimáka • Bakterie na povrchu bičíkovců z termitů • Bakterie v cytoplasmě prvoka Barbulonympha
Směry HGT • Prokaryota → Prokaryota (šíření genů rezistence na antibiotika přitáhlo pozornost k HGT) • Eukaryota → Eukaryota (méně časté; např. u hub) • Prokaryota → Eukaryota (např. endosymbiotický genový transfer) • Eukaryota → Prokaryota (zřídkavé, limitují introny; např. geny pro tubulin a aktin v bakteriích)
I. HGT Eukaryota → Prokaryota • Geny pro proteiny cytoskeletu (exkluzivně euk. geny) • Alfa a beta tubulíny, kinesin u bakterie Prostheobacter • Aktin a profilin u sinice Microcystis aeruginosa • Eukaryotické geny v genomu intracelulární bakterie Legionella (zdroj: vodní a půdní améby) • Améby jako tréninkové tábory bakt. adaptace pro euk. buňky Aktin_FITC Guljamow A et al. Curr Biol. 2007;17(17):R757-9
I. HGT Eukaryota → ProkaryotaBariéry • Introny • Příležitost (velikost polulace bakterií) • Interakce mezi bakteriemi x interakce eukaryot s bakteriemi • Konjugace je specifická pro bakterie • Eukaryoti jednoduše nemají bakteriím co nabídnout
II. HGT Eukaryota → Eukaryota • Teoreticky nejjednodušší (rozdíly v genové expresi u prokaryot a eukaryot) • Pravděpodobně podceňovaný • Častý u hub (nejvíc sekvenčních dat) • Účast virů a transpozonů
II. HGT Eukaryota → Eukaryota • Šíření virulenčních faktorů→ hospodárské ztráty • Tan spot disease pšenice • Recentní přenos genu ToxA mezi houbami Stagonospora nodurum a Pyrenophora tritici-repentis • ToxA - 99,7% identita, u příbuzných hub chybí
II. HGT Eukaryota → Eukaryota • Šíření virulenčních faktorů • Sdílení genů pro virulenci mezi houbou Nectria haematoccoca a Neocosmospora boniensis (patogeni hráchu) • Skupina genů (PEP klastr) s nižším obsahem G/C a jinou preferencí kodonů; superumerary chromosom; chybí u nepatogenních izolátů Nectria
III. HGT Prokaryota → Eukaryota • Rozsah: 0 (člověk, mnohobuněční) - stovky genů (prvoci) • U protist nejrozšířenější • Příležitost (všude přítomné bakterie) • Bakteriotrofie x eukaryotrofie (méně častá)
Zdroje genů pro eukaryota • Mechanizmus HGT u eukaryot nejasný • Organely • Endosymbionti • Potrava • Viry a transpozony
Klíčové události eukaryogenezy Vznik chloroplastu Vznik mitochondrie Archebaktérie Proteobaktérie Cyanobaktérie Timmis et al. (2004)
Cyanobaktérie: Synechocystis: 3573 kbp/ 3168 proteinů Prochlorococcus marinus: 1660 kbp/ 1884 Nostoc punctiforme: 9000 kbp/ 7400 Řasy: Porphyra purpurea: 191 kbp/ 200 Cyanidium caldarium: 165 kbp/ 197 Guillardia theta: 122 kbp/ 148 Cyanophora paradoxa: 136 kbp/ 136 Odontella sinensis: 120 kbp/ 124 Euglena gracilis: 143 kbp/ 58 α-proteobaktérie: Caulobacter crescentus: 4017 kbp/ 3767 Mesorhizobium loti: 7596 kbp/ 7281 Bradyrhizobium japonicum:9100 bp/ 8300 Prvoci: Reclinomonas americana: 69 kbp/ 67 Malawimonas jakobiformis: 47 kbp/ 49 Naegleria gruberi: 50 kbp/ 46 Dictyostelium discoideum: 56 kbp/ 40 Plasmodium falciparum: 6 kbp/ 3 !Hydrogenosomy, mitosomy: 0 kbp/ 0 Velikosti a kódovací kapacity organelových a prokaryotických genomů Timmis et al. (2004)
Organelové geny formují eukaryotické chromosomy Timmis et al. (2004)
Hlavní typy HGT u chloroplastu jádro HGT EGT (časté) plastid HGT (zřídka) Keeling PJ (2009)
Osud organelového genu Zachování obou homologů (zřídkavě). Trendem je spíše redukce organelových genomů. B. Ztráta organelového genu. Jaderný homolog kóduje produkt, který funguje v cytoplazmě i v organele. C. Přesun genu endosymbionta do hostitelského genomu a jeho společná existence s hostitelskou kopii. D. Náhrada hostitelského genu genem organelovým (tzv. endosymbiotic gene replacement). Hodně příkladů u anaerobních protist, které mají mitochondrie bez genomu. E. Přijetí genu nepříbuznéhoorganelovému endosymbiontovi (dodání nové funkce organele); přechodné endosymbiózy.
James R. Brown Nature Reviews Genetics 4, 121-132
Endosymbiotický genový transfer Je důsledkem endosymbiózy Integrace mitochondriální a plastidové DNA do eukaryotického jádra (stále aktivní). Organelová DNA má velký podíl na množství genů v eukaryotických chromozomech. Korelace množství organelové DNA v jádře a množství organel. Přemístěné geny kódují proteiny, které jsou importované do všech částí buňky, nejen zpátky do organely původu.
HGT je zdrojem nových adaptací • Ekologicky specializovaní prvoci (anaerobní paraziti, bachoroví nálevníci, extremofilové) mají hodně bakteriálních genů • HGT však nesouvisí jen s extrémními adaptacemi (půdní améby) Řasa Galdieria sulphuraria Bachorový nálevník Ophryoscolex Trichomonas vaginalis
Galdieria sulphuraria • Extremofilní červené řasa (Cyanidiophyceae) • Flexibilní metabolizmus • Jedinečná adaptace na extrémní podmínky díky HGT(pH 0-4; teplota 56°C; tolerance toxických kovů; HGT až 5% genomu) • Bakteriální geny pro různé iontové kanály • Méně intronů a vyšší GC obsah • Donoři: extremofilní bakterie a archebakterie Schönknecht et al. (2013)
Mnohobuněčné organizmy a jejich okolí poskytují habitat pro mikroby. Transfer genů má význam pro adaptaci na toto nové prostření.
HGT a kolonizace prostředí • Bakteriální β-glukuronidáza u půdních hub → využití glukuronidů z moči obratlovců jako zdroje energie • Bakteriální hydrolázy u bachorových hub → využití celulózy z bachoru býložravců • Zpracování komplexních sacharidů z bachoru býložravců – 148 prokaryotických genů u nálevníků
HGT a kolonizace prostředí • Aktinobakteriální kutináza u nepříbuzných vláknitých hub a oomycét → degradace rostlinní ochranné bariéry • HGT jako mechanizmus evoluce virulence Phytophtora (oomycéta)
HGT a proteomy parazitických protist • Převažuje HGT ve směru Prokaryota → Eukaryota • Enzymy metabolických drah (metabolizmus aminokyselin a sacharidů, ale i geny pro adaptaci k parazitizmu) • Široké spektrum prokaryotických donorů, především ale bakterie sdílející habitat s parazity
Prokaryotické geny u parazitických protist • Nejvíc recentních HGT představují single copy geny • Duplikace genů • Masivní amplifikace genových rodin – Trichomonas vaginalis (2 bakteriální geny pro hypotetické proteiny expandovali do rodin o 260 až 1200 kopií) Alsmark et al. (2013)
Prokaryotické geny u parazitických protist • Enzymy klíčových drah – 62% (metabolismus aminokyselin a sacharidů) • Až 35% geny pro hypotetické proteiny (potenciálně membránové proteiny) • Adaptivní význam pro parazitický život • Dárci genů: bakterie sdílející habitat s parazity Science 24 December 2010: vol. 330 no. 6012 1768-1773
Trichomonas, Entamoeba, Giardia • Extracelulární paraziti sliznic • HGT – zpracování sacharidů, glykanů, aminokyselin a lipidů • Adaptivní význam v prostředí! • Trichomonas vaginalis – degradace hostitelských glykanů (příklad celé metabolické dráhy získané HGT)
Trichomonas, Entamoeba, Giardia • Život v prostředí bez kyslíku • Nezískávají energii oxidativní fosforylaci ale jinými cestami (glykolýza, fermentace, substrátová fosforylace) • Využití aminokyselin jako zdroje energie Např. E. histolytica – HGT enzymů pro degradaci aminokyselin; tryptofanáza u T. vaginalis
Taxonomic distribution of putative donors of genes to Entamoeba histolyticaand Trichomonas vaginalis. Alsmark C. et al. (2009)






















