Download
1 / 32
320 likes | 609 Views
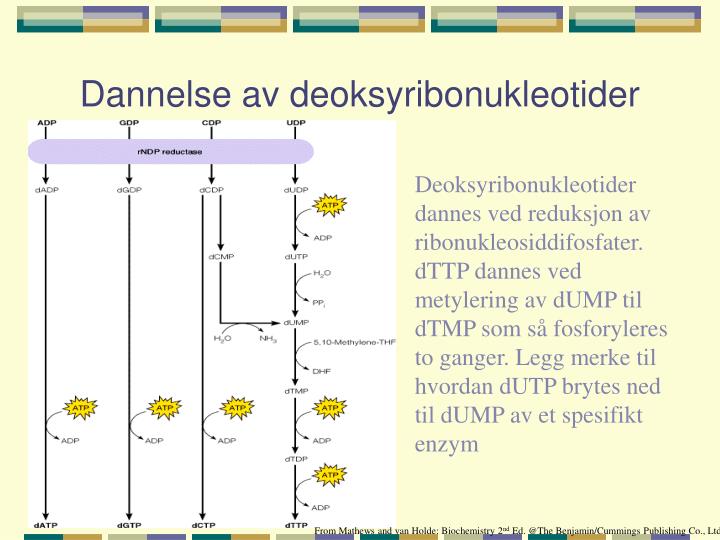
Dannelse av deoksyribonukleotider. Deoksyribonukleotider dannes ved reduksjon av ribonukleosiddifosfater. dTTP dannes ved metylering av dUMP til dTMP som så fosforyleres to ganger. Legg merke til hvordan dUTP brytes ned til dUMP av et spesifikt enzym.

E N D
Dannelse av deoksyribonukleotider Deoksyribonukleotider dannes ved reduksjon av ribonukleosiddifosfater. dTTP dannes ved metylering av dUMP til dTMP som så fosforyleres to ganger. Legg merke til hvordan dUTP brytes ned til dUMP av et spesifikt enzym From Mathews and van Holde: Biochemistry 2nd Ed. @The Benjamin/Cummings Publishing Co., Ltd
Ribonukleotid reduktase • Det finnes 4 velkarakteriserte klasser av ribonukleotid reduktaser: • Klasse I med tyrosylradikal som stabiliseres av et binukleært Fe(III)-kompleks med okso-bro. Bred fordeling blant prokaryoter og nesten alle eukaryoter • Klasse 2 med kofaktoren 5’-deoksyadenosylkobalamin (koenzym B12). Prokaryoter • Klasse 3 med [3Fe-4S]- eller [4Fe-4S]-kluster som krever SAM og NADPH for aktivitet. Anaerobe prokaryoter som ikke kan syntetisere koenzym B12 • Klasse 4: Mn(II)-holdige proteiner
Ribonukleotid reduktase Dette er klasse 1-enzymet fra E. coli. α2β2-dimer som kan oppdeles i to inaktive homodimerer, R1 (α2, 761 aa/subenhet) og R2 (β2, 375 aa/subenhet) From Mathews and van Holde: Biochemistry 2nd Ed. @The Benjamin/Cummings Publishing Co., Ltd
Dannelse av et radikal i det aktive sete i RNR Proposed electron-coupled proton-transfer pathway between the tyrosyl radical (?Tyr122) of R2 and Cys439 of R1 (a) and the reverse of this process (b)26 . Although Trp356 (shown in blue), which lies at the C-terminus of R2, is implicated in the electron-coupled proton-transfer process, the crystal structure of R2 does not include this residue, because of its disorder 3 . Electron-coupled proton transfer is formally equivalent to hydrogen-atom transfer; however, the structure suggests that direct hydrogen-atom transfer between Trp48 and Tyr122 is not feasible. Thus the electron and proton must arrive at Tyr122 by divergent pathways. In (b), conformational flexibility around Fe1, involving Glu238, His118, Asp237 and Try48 would be necessary for regeneration of the ?Tyr. Red denotes positions of coupled electron and proton transfer; ? denotes the divergence of these two processes. Fra Stubbe and Riggs-Gelasco, TIBS 23 (1998) 438-443
Ribonukleotid reduktase: en mulig mekanisme • Tyrosylradikalet mottar hydrogenatom (et elektron og et proton) fra en SH-gruppe i det aktive sete, slik at et fritt radikal dannes der • Radikalet fjerner et H-atom fra C3’ • Syrekatalysert kløyving av C2’-OH-bindingen gir et radikal-kationintermediat • Radikal-kationintermediatet reduseres av enzymets redoks-aktive sulfhydrylpar. 3’-radikalet mottar et H-atom fra proteinet og det frie radikal i proteinet gjendannes • S-S-broen reduseres av tioredoksin/glutaredoksin 1 5 2 4 3 From Mathews and van Holde: Biochemistry 2nd Ed. @The Benjamin/Cummings Publishing Co., Ltd
Reduksjon av RNR From Mathews and van Holde: Biochemistry 2nd Ed. @The Benjamin/Cummings Publishing Co., Ltd
Tymidinnukleotider I: dannelse av dUMP Deoksycytidin kinase dCMP deaminase dUTPase Tymidin kinase
Tymidinnukleotider II: omsetning av dUMP Tymidylat syntase Tymidin kinase
Gjendannelse av N5,N10-metylen-THF fra DHF Tymidylat syntase Dihydrofolat reduktase Serin hydroksymetyl-transferase
Hemming av DHF reduktase DHF-analoger som metotreksat, aminopterin og trimetoprim bindes til DHF reduktase med 1000 ganger høyere affinitet enn DHF og inhiberer enzymet. Resultatet er at HF ikke gjendannes, noe som gjør at DNA-syntese, purinsyntese og syntese av aminosyrene histidin og metionin blokkeres
Nukleotidmetabolisme: en grov oversikt Fosforibosyltransferasene er viktige for salvage av puriner. To enzymer: adenin fosforibosyl-transferase (APRT) og hypoxantin-guanin fosforibosyltransferase. Mangel på HPRT: Økt PRPP-konsentrasjon gir økt purinsyntese, økt urinsyre. Gikt, Lesch.Nyhan Nukleosidaser
GMP IMP AMP deaminase nukleotidase Guanosin Adenosin Inosin adenosin deaminase Purinnukleosid fosforylase (PNP) Guanin Hypoxantin guanin deaminase Xantin oksidase Xantin oksidase Urat Xantin Nedbrytning av puriner Adenosin og deoksyadenosin brytes ikke ned av PNP hos pattedyr. I stedet deamineres adeninnukleotider og –nuk-leosider av AMP deaminase og adenosin deaminase til de tilsvarende inosinderivatene som så nedbrytes videre
Mekanismen for xantin oksidase Det reduserte enzymet vil senere reoksideres av O2 under dannelse av H2O2
Purinnukleotidsyklus Denne syklusen spiller en viktig metabolsk rolle i skjelettmuskel-vev. Dette vevet mangler de fleste enzymene som står for anaplerotiske reaksjoner i andre vev (syntese av TCA-syklusintermediater). Fumarat fra purin-nukleotidsyklus fyller denne rollen i muskelvev
Nedbrytning av pyrimidiner Som for purinnukleotider degraderes pyrimidin-nukleotidene til baser ved defosforylering, deaminering og bryting av glykosidbindinger. Basene som dannes, uracil og tymin, nedbrytes videre ved reduksjon, ikke ved oksidasjon som for purinene
Disorder Defect Nature of Defect Comments Gout PRPP synthetase increased enzyme activity due to elevated Vmax hyperuricemia Gout PRPP synthetase enzyme is resistant to feed-back inhibition hyperuricemia Gout PRPP synthetase enzyme has increased affinity for ribose-5-phosphate (lowered Km) hyperuricemia Gout PRPP amidotransferase loss of feed-back inhibition of enzyme hyperuricemia Gout HGPRTa partially defective enzyme hyperuricemia Lesch-Nyhan syndrome HGPRT lack of enzyme Gout and severe malfunction of the nervous system. Selfmutilation SCID ADAb lack of enzyme B and T lymphocytes. RNR Immunodeficiency PNPc lack of enzyme see above Renal lithiasis APRTd lack of enzyme 2,8-dihydroxyadenine renal lithiasis Xanthinuria Xanthine oxidase lack of enzyme hypouricemia and xanthine renal lithiasis von Gierke's disease Glucose-6-phosphatase enzyme deficiency see above Sykdommer forbundet med purinmetabolismen
Disorder Defective Enzyme Comments Orotic aciduria Type I orotate phosphoribosyl transferase and OMP decarboxylase retarded growth and severe anemia Orotic aciduria Type II OMP decarboxylase see above Orotic aciduria (mild, no hematologic component) the urea cycle enzyme ornithine transcarbamoylase, is deficient increased mitochondrial carbamoyl phosphate exits and augments pyrimidine biosynthesis; hepatic encephalopathy b-aminoisobutyric aciduria Transaminase benign, frequent in Orientals Drug induced orotic aciduria OMP decarboxylase Allopurinol and 6-azauridine treatments cause orotic acidurias; their catabolic by-products inhibit OMP decarboxylase Sykdommer forbundet med pyrimidinmetabolismen
Syntese av NAD(P)+ Nikotinsyre og nikotinamid får overført ribosefosfat fra PRPP. Nikotinsyremonofosfat kan også dannes fra histidin via kinolinsyre. AMP tilkobles for dannelse av dinukleotidene (merk pyrofosfatbindingen!), nikotinsyre adenin dinukleotid amideres til NAD+, og NAD+ fosforyleres i 2’-stilling i ribosen som er knyttet til nikotinamid for dannese av NADP+
Syntese av FMN og FAD ATP Riboflavin fra kosten fosforyleres til FMN. AMP kobles på for dannelse av FAD
Syntese av CoA Pantotensyre består av pantoinsyre bundet til β-alanin via en amid(peptid)-binding, og kan ikke syntetiseres av mennesker. Pantotensyre fosforyleres, en ny peptidbinding dannes mellom pantotensyre og cystein, produktet dekarboksyleres til 4’fosfopantetein, AMP henges på og ribosen fosforyleres i 3’-stilling for dannelse av CoA
Baser, nukleosider, nukleotider Syrelabil N-glykosidbinding, syrelabil dersom basen er et purin
Struktur av nukleinsyrer: RNA Til venstre en vanlig strukturformel, over en mer skjematisk fremstilling. Sekvensen kan også skrives 5’-ApUpCpGp ApUpCpGp AUCG
Hydrolyse av RNA under alkaliske betingelser Hydroksylgruppen i 2’-stilling angriper fosfatgruppen bundet i nabostillingen (3’). Det dannes en syklisk diester som vil hydrolyseres, enten til et nukleosid-3’-fosfat eller et nukleosid-2’-fosfat






















