Download
1 / 41
410 likes | 556 Views
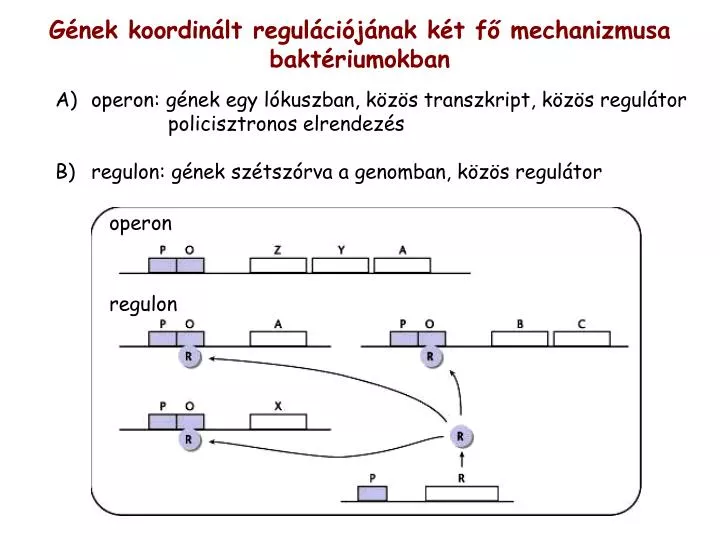
operon. regulon. Gének koordinált regulációjának két fő mechanizmusa baktériumokban. operon: gének egy lókuszban, közös transzkript, közös regulátor policisztronos elrendez és regulon: gének szétszórva a genomban, közös regulátor. negatív szabályozás. pozitív szabályozás. indukció.

E N D
operon regulon Gének koordinált regulációjának két fő mechanizmusabaktériumokban • operon: gének egy lókuszban, közös transzkript, közös regulátor policisztronos elrendezés • regulon: gének szétszórva a genomban, közös regulátor
negatív szabályozás pozitív szabályozás indukció KATABOLIKUS KATABOLIKUS indukálószer inaktív aktivátor indukálószer represszor inaktív represszor aktív aktivátor represszált derepresszált inaktív RNS polimeráz RNS polimeráz represszió BIOSZINTETIKUS BIOSZINTETIKUS indukálószer indukálószer inaktív represszor aktív aktivátor aktív represszor inaktív aktivátor represszált inaktív derepresszált indukált RNS polimeráz RNS polimeráz A TRANSZKRIPCIÓS SZABÁLYOZÁS FŐBB GLOBÁLIS STRATÉGIÁI PROKARIÓTÁKBAN
A transzkripciós faktorok és aDNS közötti specifikus kölcsönhatásún. Hélix-Turn-Hélix (HTH) motívumon megy keresztül HTH Motívumok: csgD: NNEIARSLFISENTVKTH LY merR: IGEVALLCDINPVTLRAWQR luxR: SWDISKILGCSERTVTFHLT lehet a faktor N vagy C terminálisán, a másik végen szokott lenni a ligand, kofaktor kötő régió
BAKTERIÁLIS TRANSZKRIPCIÓS FAKTOROK FŐBB CSALÁDJAI Faktor család Tagok AraC család AraC, MelR, RhaS, RhaR, SoxS LysR család LysR, OxyR, MetR, CysB Crp család Crp, Fnr MerR család SoxR Két komponensű NarL, OmpR, Arc szabályozó család Lac represszor család LacI, GalR MetJ család MetJ
Aktiváció a gén expresszióban I. Kölcsönhatás: - CTD-nel (CRP) - 70 4-es régiójával ( cI aktivátor) - NTD-nel (CRP) - alegységgel (DnaA) - ’ alegységgel (N4 single-stranded DNA kötő fehérje) - CTD-nel és 70 4-es régiójával (FNR) Positive activation of gene expression Virgil A Rhodius, Stephen JW Busby Current Opinion in Microbiology 1998, 1:152-159.
Aktiváció a gén expresszióban II. Promóter konformáció megváltoztatása: - “-35” és “-10” régió azonos oldalra kerül (MerR, SoxR) - DNS visszahajlik és az aktiváló cisszekvencia RNAP fölé kerül - DNS konformáció változást indukál (FIS, IHF) Positive activation of gene expression Virgil A Rhodius, Stephen JW Busby Current Opinion in Microbiology 1998, 1:152-159.
Távoli aktivátor helyek segítséget igényelnek DNS-hajlítófehérje (pl. IHF) Specifiuskötőhely
Positive activation of gene expression Virgil A Rhodius, Stephen JW Busby Current Opinion in Microbiology 1998, 1:152-159 Aktiváció a gén expresszióban III. 2 aktivátortól függő promóterek: - az aktivátor kötődése egy másik aktivátortól függ (eukarióta) - aktivátor kötődése egy másik aktivátor áthelyeződését eredményezi (CRP-MalT a malK promóteren) - független aktiváció (70 vagy NTD és CTD) - represszor müködését gátolja (FIS-NARL/P-FNR)
FNR - fumarát nitrát reduktáz regulátor - citoplazmatikus szenzor-regulátor - dimer[4Fe-4S]2+ DNS-t köt - monomer[2Fe-2S]2+ inaktív - aenaerob respirációra (+) vagy (-) hatás - pO2 1 mbar alatt - TTGAT-N4-ATCAA konszenzus szekvencia - [2Fe-2S]2+ [4Fe-4S]2+ (in vitro) Cys, Fe, DTT, NifS - Pseudomonas: ANR; Bacillus: FNR Rhodobacter sphaeroides: FnrL O2 FNRox FNRred
Két komponensű szabályozó rendszerek Komponensek - egy szenzor kináz és egy DNS kötő regulátor - E. coli genom 2% - kb 50 különböző 2 komponensű rendszer - 3 alcsalád: OmpR, FixJ és NtrC
szenzor kináz fehérje DNS kötő fehérje szignál Érzékelő Foszforilációs P P DNS Felvevő Felvevő DNS kötő DNS kötő transzfoszforiláció DNS Érzékelő Érzékelő Foszforilációs Foszforilációs A bakteriális kétkomponensű szabályozó rendszerek működése elve
OmpR - OmpR (E. coli): porin szerveződés szabályozása ozmózis változás hatására - általában 70 használó transzkripciót aktivál - kölcsönhatás az RNS polimeráz alegységének C terminálisával - ha az N terminális foszforilálódik megszünik a gátló hatása a C terminális DNS kötő domén felé
FixJ - általában 70 használó transzkripciót aktivál - receiver domén deléciója esetén aktív transzkripció lesz NtrC - N terminális receiver és C terminális DNS kötő domén között egy központi ATPáz domén (glicin gazdag “Walker box”) - DNS kötő domén FIS-hez homológ (FIS: eukarióta enhancer kötő fehérje) - általában 54 használó transzkripciót aktivál
P ArcA/B - aerobic respiratory control - ArcB (szenzor kináz): sejt redox és metabolikus helyzet (elektron transzport változást érzékel) - ArcA(citoplazmatikus regulátor): ArcB foszforilálja aktív - pO2 1-5 mbar között - TATTTaa konszenzus szekvencia - Haemophilus: ArcA - E. coli homológ gén: OmpR ArcA ArcB O2 P ArcAP
P oxigén mentes környezet ArcB FNR ArcA Az ArcAB és FNR anaerob aktivációja
NarX NO3 NarLP P NarQ NarPP NarX/L és NarP/Q - nitrát regulator - NarX és NarQ: membrán szenzor kináz - NarL és NarP: citoplazmatikus regulátor - szignál: nitrát és nitrit - nitrát metabolizmusra hat - NarL és NarP különböző génekre különböző atás a génexpresszió finom hangolása Kétkomponensű rendszerekvége
A lac operon kettős szabályozása • laktóz (allolaktóz) indukál • glükóz gátol, cAMP/CAP-n • keresztül • glükóz/egyéb cukor kiiktatása tápból nem célszerű • glükóz szabályozás kikapcsolása
UV lacUV5 lac promóter lac (és trp) alapú promóterek erősség “-35” “-10” trp AATGAGCTGTTGACAATTAATCATCGAACTAGTTAACTAGTACGCA tac AATGAGCTGTTGACAATTAATCATCCGGCTCGTATAATGTGTGGA lacUV5 CCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATAATGTGTGGA lac CCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATGGTGTGTGGA
AraC Ara C PBAD • Ara PBAD represszió • +Ara PBADindukció AraC RNS polimeráz PBAD A transzkripciós faktorok sokoldalúak….
a) Rho kötődik és üldözi a polimerázt a) hurok képződés indul b) hurok képződés, polimeráz megáll b) hurok képződés indul hibrid destabilizálódik c) termináció c) Rho helikáz felszabadítja a transzkriptet, termináció Transzkripció termináció baktériumokban Rho független Rho függő
A transzkripció és a transzláció párhuzamosanmegy baktériumokban
A triptofán operon szerkezete protein antranilát szintáz indol-glicerin szintáz triptofán szintáz
Termináció - antitermináció túl sokat nem lehet vele kezdeni, génen belüli sajátság
mRNS degradáció baktériumokban mRNS stabilitás prokariótákban néhány perc, eukariótákban órás nagyságrend előbb utóbb minden RNS lebomlik mRNS stabilitását meghatározó faktorok: - belső, saját szerkezet - a környezet hatására bekövetkezett változás a degradációs apparátusban puf operon (a fotoszintetikus komplex komponensei) Rhodobacter capsulatus degradációja O2 hatására felgyorsul policisztronos rendszerek esetén az alegységek arányának szabályozása a mRNS régióinak eltérő stabilitásával
mRNS degradáció baktériumokban, vizsgálati módszerek • - transzkripció gátlása (pl. rifampicin) t=0 időpontban, • majd időközönként mintavétel és RNS analízis (Northern..) • a degradációs mechanizmusban szerepet játszó gének deléciója, • hőmérséklet érzékeny expressziós változatának kialakítása • - in vitro transzkripció jelölt nukleotidokkal, a kapott termék inkubációja a sejtextraktummal különböző ideig, majd analízis gélelektroforézissel, kvantitálás
RNázok, RNS degradáció Bacterial exoribonucleases Polynucleotide phosphorylase Ribonuclease PH Ribonuclease II Ribonuclease R RNase D Ribonuclease T Ribonuclease BN Oligoribonuclease Bacterial endoribonucleases Ribonuclease I Ribonuclease III Ribonuclease P Ribonuclease E Ribonuclease HI A degradáció iránya virtuálisan 5' 3' irányú, de ilyen enzimaktivitást nem lehet kimutatni prokariótákban Megoldás: kombinált enzimműködés degradoszóma
Az 5’ végi struktúra stabilizáló hatása • a stabillizálódás a mRNS hurok struktúrájában van • nem a riboszóma véd, • a stabilizáló effektus átvihető más génekre
mRNS-eket stabilizáló (védő) tényezők • 5’ végi trifoszfát • RNS struktúra • riboszóma
Az RNaseE elsődleges felépítése Érdekes módon sok bakteriális genomban nincs meg
A degradoszóma komponenseiI. Endoribonuclease E (RNáz E) • 1061 aminosav 118 kDa fehérje, virtuálisan 180 kDa(oka prolin gazdag régió) • felismerő hely: (A/G)AUU(A/T) vagy egy komplex másodlagos struktúra • 5' monofoszfátot preferál 5' trifoszfát stabilizál N-terminális régió (50 kDa) hasonlít a Caf-re (cytoplasmic axial filament protein) feltételezett funkció a belső RNS transzportban N terminális 70 aa (S1 domén) hasonlít a PNPase és RnaseII (illetve (CSP, cold shock protein, RNS chaperon) RNS kötő doménjére C-terminálisa degradoszóma egyéb komponenseire megfelelő kötő domének
A degradoszóma komponenseiII. • PNPase (polynucleotide phosphorylase) • 78 kDa alegységek, homotrimer • 3' 5' foszfát függő processzív exonukleáz, • ribonukleotid difoszfátok képződnek • poliadenilációs aktivitás • Polyphosphate Kinase (PPK) • funkció: ATP regeneráció, polifoszfát (inhibiálja a degradációt) eltávolítás • ppk mínusz törzs : megnövekedett mRNS stabilitás • 80 kDa alegységek, homotetramer, sok van E. coli-ban
ATP hiányában a hurokstruktúra stabil marad A degradoszóma komponenseiIII. • Helikáz • ATP függő DEAD (Asp-Glu-Ala-Asp) helikáz • 50 kDa RhlB • a másodlagos struktúrák kinyitása szétroncsolása
Egyéb – mRNS degradációjában résztvevő – enzimek • RNáz II • 70 kDa monomer, • a sejt 3' 5' exoribonukláz aktivitásának 90%-a • ribonukleotid monofoszfátok képződnek • a PNPáz-zal együttes deléciója letális !!! • PolyA polimerázok • PAPI 53 kDa • PAPII 35 kDa • poliadeniláció, 15- 50 bázis hosszú • mRNS instabilitás